За счет чего сокращается скелетная мышца: МЕХАНИЗМ СОКРАЩЕНИЯ СКЕЛЕТНЫХ МЫШЦ | Библиотека тренера
МЕХАНИЗМ СОКРАЩЕНИЯ СКЕЛЕТНЫХ МЫШЦ | Библиотека тренера
+7 (914) 798 17 43
Назад к списку материалов
Никита Ющенко (автор)
ВОЗБУЖДЕНИЕ МЕМБРАНЫ МЫШЕЧНОГО ВОЛОКНА
Рис. Нервно-мышечный синапс
Нервы, подходящие к мышцам, содержат три вида волокон:
- чувствительные;
- двигательные;
- вегетативные (иннервируют мышечные сосуды).
Окончания двигательных нейронов (мотонейронов) подходят к каждому мышечному волокну. Между аксоном мотонейрона и волокном находится нервно-мышечный синапс.
- Когда мышца расслаблена, в саркомерах нити актина лишь частично заходят в просветы между нитями миозина.
 Актин блокирован другими белками, и миозин не может с ними взаимодействовать.
Актин блокирован другими белками, и миозин не может с ними взаимодействовать. - Когда по мотонейрону проходит импульс из ЦНС, в нервно-мышечный синапс выделятся нейромедиатор ацетилхолин и связывается с рецепторами на мембране мышечного волокна. В ответ на это в мышечное волокно входят ионы натрия, заряд мембраны меняется, и она переходит в возбужденное состояние.
 Актин блокирован другими белками, и миозин не может с ними взаимодействовать.
Актин блокирован другими белками, и миозин не может с ними взаимодействовать. ПЕРЕДАЧА ВОЗБУЖДЕНИЯ НА МЕМБРАНЫ СПС. СОКРАЩЕНИЕ МИОФИЛАМЕНТОВ
Передача возбуждения на мембраны СПС
Рис. Механизм сокращения мышечных волокон
Далее возбуждение распространяются по Т-трубочкам вглубь клетки, переходит на мембраны саркоплазматической сети, и из цистерн СПС высвобождаются ионы кальция.
- Ионы кальция связываются с белками, блокирующими актин; конфигурация белковых молекул меняется, и миозин получает возможность соединиться с актином. Это активирует фермент АТФ-азу миозина, который входит в состав миозиновых головок.

Сокращение миофиламентов
Рис. Взаимное расположение миофиламентов в расслабленном и сокращенном мышечном волокнеАТФ-аза миозина расщепляет АТФ, высвобождая энергию для сокращения. За счет этой энергии головки миозина тянут актиновые нити, продвигая их к центру саркомера, и саркомер сокращается (укорачивается).
- Затем, также за счет энергии АТФ, кальций перемещается обратно в цистерны, а головки миозины отделяются от актина. Нити возвращаются в исходное положение — миофибрилла расслабляется (удлиняется).
ИСТОЧНИКИ:
- Основы анатомии, физиологии и биомеханики : учебник / М.Б. Андреева, Л.А. Белицкая, В.А. Меркурьев; под ред. Д. Г. Калашникова. — М.: Практическая медицина, 2019. — 336 с.
- Анатомия человека (с основами динамической и спортивной морфологии): Учебник для институтов физической культуры. — Изд. 14-е. / Под. ред. Б. А. Никитюка, АА. Гладышевой, В. Ф. Судзиловского. — М.: Спорт, 2018. — 624 с., ил.
- Физиология человека: учебник для вузов физической культуры/В.И. Тихоревский: «Физкультура, образование и наука», 2001.
 — Изд. 14-е. / Под. ред. Б. А. Никитюка, АА. Гладышевой, В. Ф. Судзиловского. — М.: Спорт, 2018. — 624 с., ил.
— Изд. 14-е. / Под. ред. Б. А. Никитюка, АА. Гладышевой, В. Ф. Судзиловского. — М.: Спорт, 2018. — 624 с., ил.Поделиться
Share
Поделиться
Поделиться
Биомеханика
НЕ ЛУЧШИЕ УПРАЖНЕНИЯ | ЖИМ ШТАНГИ ИЗ-ЗА ГОЛОВЫ
Биомеханика
НЕ ЛУЧШИЕ УПРАЖНЕНИЯ | ГИПЕРЭКСТЕНЗИЯ С КРУГЛОЙ СПИНОЙ
Сокращение скелетных мышц человека
Описан процесс сокращения скелетных мышц человека, который состоит из ряда этапов.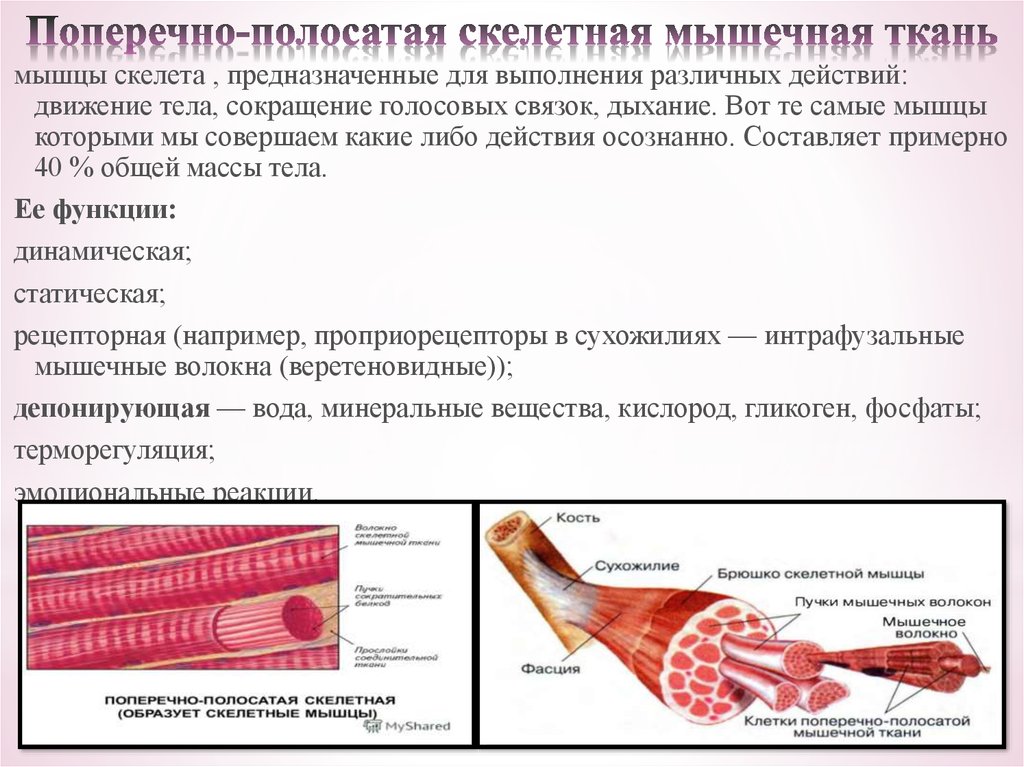 Первый этап связан с поступлением нервного импульса по аксону мотонейрона к мышечному волокну. Второй этап заключается в возникновении потенциала действия и распространении его вдоль мышечного волокна. В результате третьего этапа в саркоплазму выделяются ионы кальция, что приводит к началу взаимодействия толстого и тонкого филаментов. Четвертый этап представляет собой скольжение тонких филаментов относительно толстых, что составляет собственно сокращение мышцы.
Первый этап связан с поступлением нервного импульса по аксону мотонейрона к мышечному волокну. Второй этап заключается в возникновении потенциала действия и распространении его вдоль мышечного волокна. В результате третьего этапа в саркоплазму выделяются ионы кальция, что приводит к началу взаимодействия толстого и тонкого филаментов. Четвертый этап представляет собой скольжение тонких филаментов относительно толстых, что составляет собственно сокращение мышцы.
Давайте теперь разберемся в механизме сокращения мышцы, точнее в механизме сокращения мышечных волокон, а еще более точно в механизме сокращения миофибрилл или другими словами, в механизме сокращения саркомера. Этот процесс можно условно разделить на несколько этапов.
Поступление нервного импульса к мышечному волокну
Чтобы сократиться, мышца должна получить сигнал из центральной нервной системы (ЦНС). Такими сигналами являются импульсы, поступающие по мотонейрону к мышце.
Более подробно строение и функции мышц описаны в моих книгах:
- Гипертрофия скелетных мышц человека
- Биомеханика опорно-двигательного аппарата человека
Вспомним, что при подходе к мышце аксон мотонейрона ветвится, то есть пускает веточки к мышечным волокнам.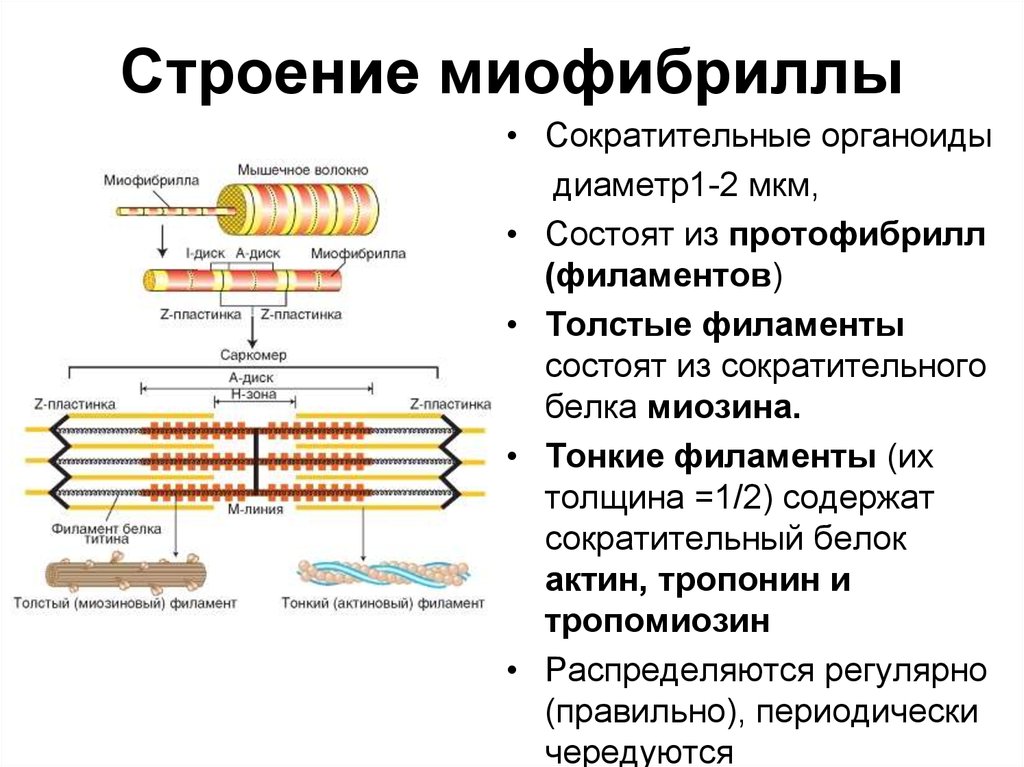 Если такого соединения нет, мышечное волокно сокращаться не будет и постепенно атрофируется.
Если такого соединения нет, мышечное волокно сокращаться не будет и постепенно атрофируется.
Возникновение потенциала действия
После того, как по аксону мотонейрона к мышечным волокнам приходит импульс, из него в области соединения выделяется ацетилхолин. Выделение этого нейромедиатора (ацетилхолина) приводит к протеканию ряда процессов, в результате которых меняется полярность сарколеммы мышечного волокна. Это называется деполяризацией сарколеммы мышечного волокна. В результате развивается потенциал действия.
Выделение ионов кальция
Потенциал действия через отверстия в сарколемме «проникает» внутрь мышечного волокна и через Т-трубочки достигает саркоплазматического ретикулума (то есть происходит деполяризация не только мембраны мышечного волокна, но и мембран Т-трубочек и саркоплазматического ретикулума). Это в конечном счете приводит к выделению из саркоплазматического ретикулума ионов кальция в саркоплазму мышечного волокна (рис. 1).
1).
Рис.1. Механизм сокращения скелетных мышц
Затем ионы кальция соединяются с тропонином (тропонин – один из белков тонкого филамента). Этот белок имеет шарообразную форму и расположен в тонком филаменте регулярно через определенные расстояния. После соединения с ионами кальция, тропонин меняет свою конфигурацию и приподнимает длинные тропомиозиновые трубки. Когда мышца неактивна, длинные трубки белка тропомиозина закрывают активные центры на актине. После того как тропомиозиновые трубки приподнимаются, на актине открываются активные центры. К ним теперь могут прикрепляться миозиновые головки.
Сокращение саркомера (гребковая гипотеза, теория скользящих нитей)
Когда миозиновая головка толстого филамента прикрепляется к тонкому филаменту, между толстым и тонким филаментами начинается взаимодействия (говорят: «Образуется поперечный мостик» (рис. 2). При взаимодействии с актином каждая миозиновая молекула ежесекундно расщепляет с выделением энергии до 10 молекул АТФ.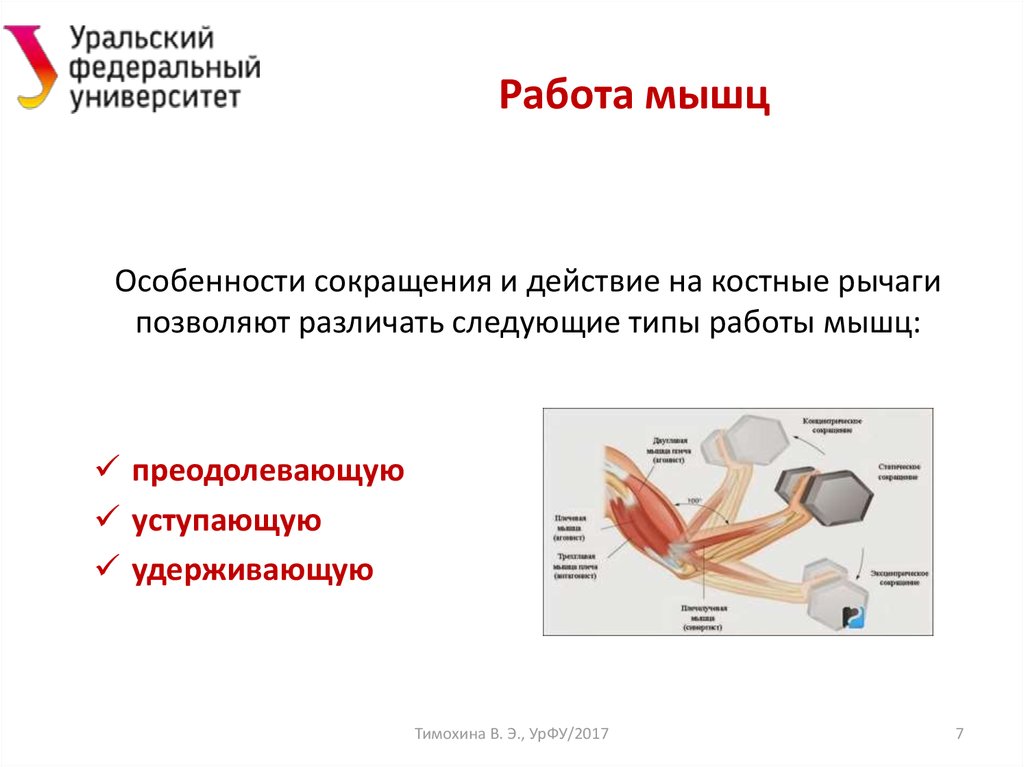 За счет энергии, высвобождающейся при расщеплении АТФ, миозиновая головка поворачивается и тянет тонкий филамент в направлении центра саркомера. Это приводит к скольжению толстого и тонкого филаментов относительно друг друга. В конце гребка (поворота) к миозиновой головке присоединяется новая молекула АТФ, что приводит к отделению головки от актина и присоединению её к новому активному участку тонкого филамента. Многократное повторение этого процесса приводит к тому, что расстояние между Z-дисками уменьшается. Следовательно, происходит уменьшение длины саркомера. Одновременное сокращение всех саркомеров, расположенных последовательно вдоль миофибриллы приводит к уменьшению её длины, длины мышечного волокна и всей мышцы в целом. Мышца работает в преодолевающем режиме.
За счет энергии, высвобождающейся при расщеплении АТФ, миозиновая головка поворачивается и тянет тонкий филамент в направлении центра саркомера. Это приводит к скольжению толстого и тонкого филаментов относительно друг друга. В конце гребка (поворота) к миозиновой головке присоединяется новая молекула АТФ, что приводит к отделению головки от актина и присоединению её к новому активному участку тонкого филамента. Многократное повторение этого процесса приводит к тому, что расстояние между Z-дисками уменьшается. Следовательно, происходит уменьшение длины саркомера. Одновременное сокращение всех саркомеров, расположенных последовательно вдоль миофибриллы приводит к уменьшению её длины, длины мышечного волокна и всей мышцы в целом. Мышца работает в преодолевающем режиме.
Прекращение импульсов, поступающих от мотонейрона к мышечному волокну приводит к расслаблению мышцы.
Рис.2. Схема, иллюстрирующая взаимодействие толстого и тонкого филаментов (Л. Страйер, 1985)
С уважением, А.В.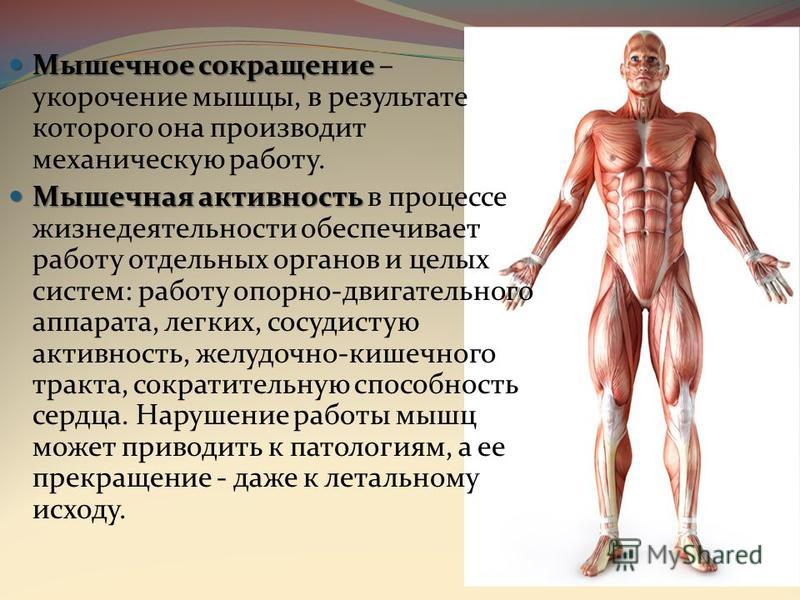 Самсонова
Самсонова
Похожие записи:
Сила тяжести
Дано определение силы тяжести. Показано, что сила тяжести является частным случаем силы гравитации. Описаны факторы, определяющие силу тяжести:…
Сила
Дано определение силы в механике. Описаны факторы, определяющие действие на тело силы: направление, точка приложения и численное значение.
Звенья тела человека как рычаги
Дано описание опорно-двигательного аппарата (ОДА) человека как системы рычагов. Приведен пример расчета силы двуглавой мышцы плеча…
Способ определения положения центров масс (центров тяжести) звеньев тела человека
Описан способ определения положения центров масс (центров тяжести, ЦТ) звеньев тела человека по Вильгельму Брауне и…
Распределение масс в теле человека
Описаны особенности распределения масс в теле человека. Дано понятие геометрии масс тела человека. Показано, что на…
Центр масс и центр тяжести тела
Описаны: центр масс (ЦМ) и центр тяжести (ЦТ) твердого тела.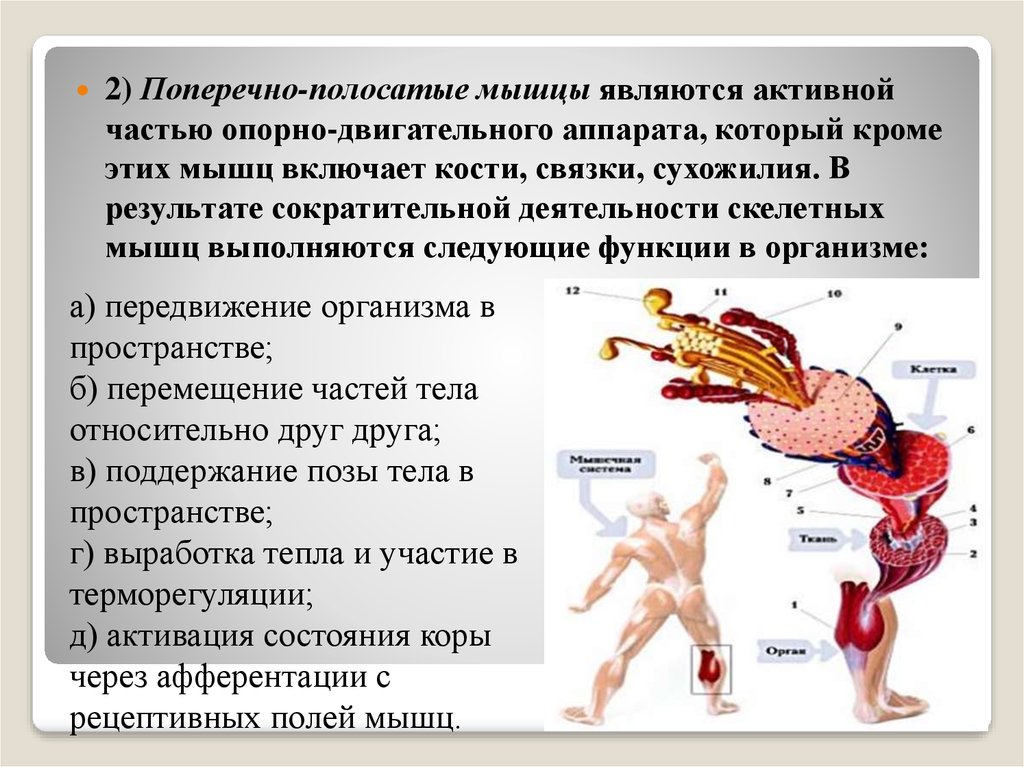 Приведены различные определения ЦМ и ЦТ тела. Показано…
Приведены различные определения ЦМ и ЦТ тела. Показано…
Метаболический стресс. Накопление лактата в мышцах
Описан механизм влияния метаболического стресса (накопления лактата) на гипертрофию мышечных волокон. Показано, что накопление лактата приводит к…
Механическое повреждение мышечных волокон
Описаны механизмы механического повреждения мышечных волокон при силовой тренировке, приводящие к гипертрофии скелетных мышц. Показано, что…
Механическое напряжение (механотрансдукция) в скелетных мышцах
Описаны процессы передачи механического напряжения в скелетных мышцах. Показано, что механическое напряжение, возникающее вследствие сокращения скелетных…
Физиология, сокращение скелетных мышц — StatPearls
Введение
Как органы, содержащие клетки, способные сокращаться, мышцы могут генерировать силу и движение. Скелетные мышцы работают вместе с костями скелета, создавая движения тела. Кроме того, он также связан с мышцами диафрагмы, пищевода и глаз. Таким образом, скелетные мышцы служат множеству целей, включая движение тела, дыхание и глотание. В отличие от гладких мышц и сердечной мышцы, скелетные мышцы сокращаются в основном в ответ на произвольные стимулы.
Таким образом, скелетные мышцы служат множеству целей, включая движение тела, дыхание и глотание. В отличие от гладких мышц и сердечной мышцы, скелетные мышцы сокращаются в основном в ответ на произвольные стимулы.
Клеточный
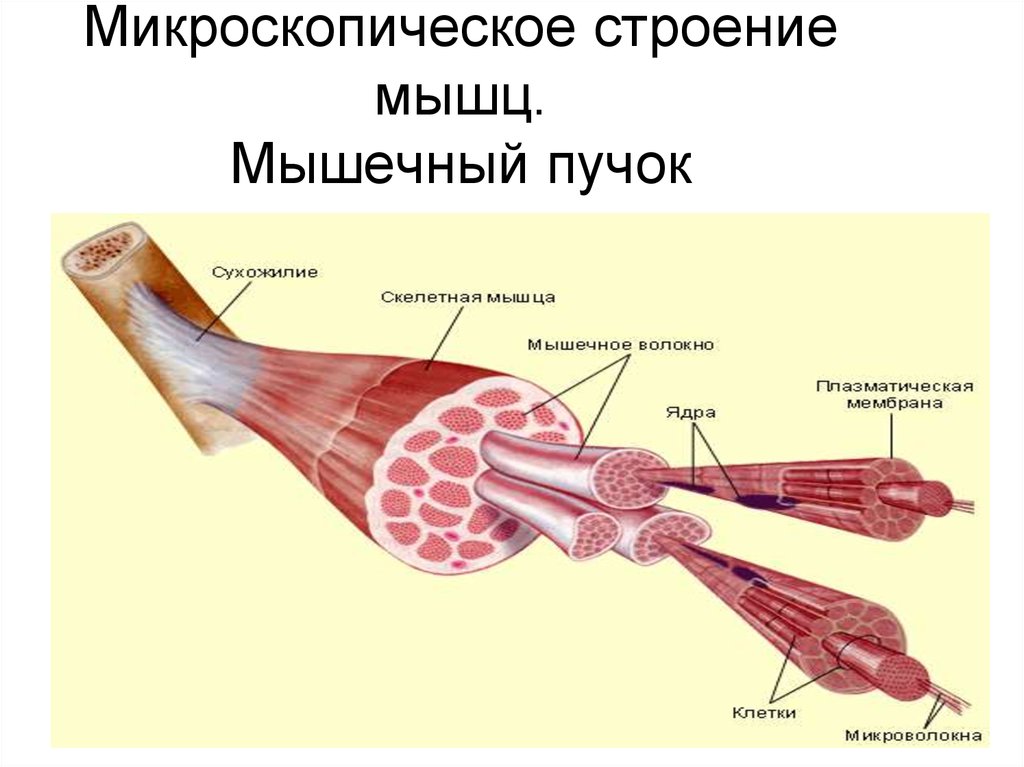
Скелетная мышца состоит из клеток, которые в совокупности называются мышечными волокнами. Каждое мышечное волокно многоядерное, его ядра расположены по периферии волокна. Каждое мышечное волокно далее подразделяется на миофибриллы, которые являются основными единицами мышечного волокна. Эти миофибриллы окружены мышечной клеточной мембраной (сарколеммой), которая образует внутри миофибриллы глубокие впячивания, называемые поперечными трубочками (Т-трубочками). Каждая миофибрилла содержит сократительные белки, описываемые как толстые и тонкие филаменты, которые расположены продольно в единицы, называемые саркомерами.
Основной единицей толстого филамента является большой белок миозин, который образован двумя парами легких цепей и одной парой тяжелых цепей. Две тяжелые цепи миозина закручиваются друг вокруг друга, образуя спиральный хвост миозина, тогда как легкие цепи взаимодействуют с тяжелыми цепями, образуя две головки миозина на другом конце. На головках находится важный участок связывания, который облегчает взаимодействие миозина с актином, белком, принадлежащим тонкому филаменту.[1]
Две тяжелые цепи миозина закручиваются друг вокруг друга, образуя спиральный хвост миозина, тогда как легкие цепи взаимодействуют с тяжелыми цепями, образуя две головки миозина на другом конце. На головках находится важный участок связывания, который облегчает взаимодействие миозина с актином, белком, принадлежащим тонкому филаменту.[1]
Другой сократительной нитью в миофибриллах является тонкая нить, в основном состоящая из трех белков: актина, тропомиозина и тропонина. Мономерная глобулярная форма актина, называемая G-актином, полимеризуется в две нити, которые скручиваются и переплетаются друг с другом, образуя нитевидный актин, называемый F-актином. По всей длине F-актина находятся сайты связывания миозина, которые скрыты нитчатым белком тропомиозином. Функция тропомиозина заключается в предотвращении взаимодействия актина и миозина, когда мышца находится в состоянии покоя, что, следовательно, предотвращает сокращение мышцы. Тропонин представляет собой комплекс из трех белков, расположенных вдоль тропомиозиновых филаментов. Первый белок, тропонин Т, способствует связыванию тропонина с тропомиозином. Тропонин I служит той же цели, что и тропомиозин, останавливая актин-миозиновое взаимодействие, блокируя сайты связывания миозина. Наконец, тропонин С связывает кальций, чтобы инициировать мышечное сокращение.[2]
Первый белок, тропонин Т, способствует связыванию тропонина с тропомиозином. Тропонин I служит той же цели, что и тропомиозин, останавливая актин-миозиновое взаимодействие, блокируя сайты связывания миозина. Наконец, тропонин С связывает кальций, чтобы инициировать мышечное сокращение.[2]
Как упоминалось ранее, толстые и тонкие нити миофибрилл организованы в единицы, называемые саркомерами. Саркомер является основной сократительной единицей миофибриллы. Z-линии разделяют каждый саркомер. Полосы А, расположенные в центре каждого саркомера, содержат толстые филаменты, которые могут перекрываться с тонкими филаментами. Полоса А далее делится на зону Н, которая не содержит тонких нитей. Выступающая линия М делит пополам зону Н и служит для соединения средних частей толстых нитей. По обеим сторонам полосы A расположены полосы I, которые содержат как тонкие нити, так и линию Z, которая проходит по середине каждой полосы I.
Механизм
Нервы, отвечающие за иннервацию мышечных волокон, называются мотонейронами. Отдельный мотонейрон и мышечные волокна, которые он иннервирует, в совокупности называются двигательной единицей. Количество мышечных волокон в двигательной единице предсказуемо зависит от функции мышцы. Например, двигательные единицы, ответственные за мимические мышцы, включают в себя значительно меньше мышечных волокон, чем двигательные единицы, отвечающие за мышцы, участвующие в таких действиях, как плавание.
Отдельный мотонейрон и мышечные волокна, которые он иннервирует, в совокупности называются двигательной единицей. Количество мышечных волокон в двигательной единице предсказуемо зависит от функции мышцы. Например, двигательные единицы, ответственные за мимические мышцы, включают в себя значительно меньше мышечных волокон, чем двигательные единицы, отвечающие за мышцы, участвующие в таких действиях, как плавание.
Сокращение скелетных мышц начинается сначала в нервно-мышечном соединении, которое является синапсом между мотонейроном и мышечным волокном. Распространение потенциалов действия на мотонейрон и последующая деполяризация приводят к открытию потенциалзависимых кальциевых (Са2+) каналов пресинаптической мембраны. Поток Са2+ внутрь вызывает высвобождение ацетилхолина (АХ) в нервно-мышечном соединении, который диффундирует к постсинаптической мембране мышечного волокна. Постсинаптическая мембрана мышечного волокна также известна как двигательная замыкательная пластинка. АХ связывается с никотиновыми рецепторами, расположенными на двигательной концевой пластинке, деполяризуя ее, что инициирует потенциалы действия в мышечном волокне.
Связь возбуждения и сокращения относится к механизму, который преобразует упомянутые выше потенциалы действия в мышечных волокнах в сокращение мышечных волокон. Потенциалы действия на мембране мышечных клеток, окружающих миофибриллы, проходят в Т-трубочки, которые отвечают за распространение потенциала действия с поверхности внутрь мышечного волокна. Т-трубочки содержат дигидропиридиновые рецепторы, которые прилегают к терминальным цистернам саркоплазматического ретикулума мышечного волокна. Когда Т-трубочки деполяризуются, их дигидропиридиновые рецепторы претерпевают конформационные изменения, которые механически взаимодействуют с рианодиновыми рецепторами на саркоплазматическом ретикулуме. Это взаимодействие открывает рианодиновые рецепторы, вызывая высвобождение Ca2+ из саркоплазматического ретикулума. В результате повышенный внутриклеточный Са2+ прикрепляется к тропонину С тропонинового комплекса на тонких филаментах. Взаимодействие между Ca2+ и тропонином C проявляет кооперативность, что означает, что каждый Ca2+, который связывает тропонин C, увеличивает сродство связывания тропонина C со следующей молекулой Ca2+, всего до четырех ионов Ca2+ на тропонин C.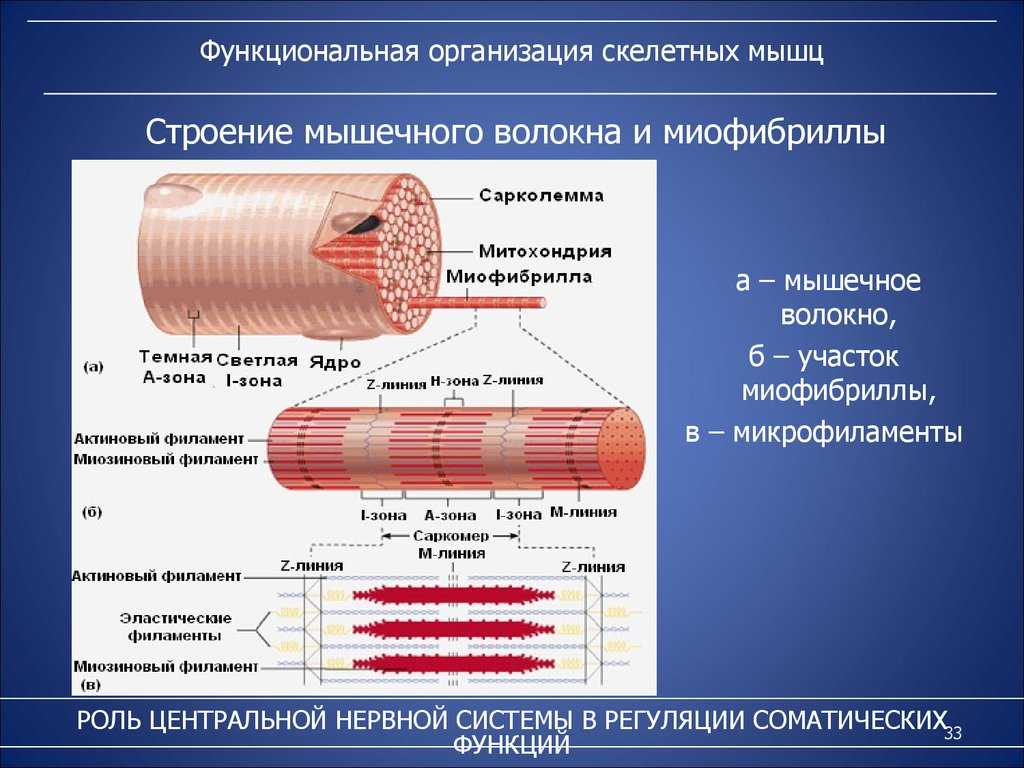 В результате связывания Ca2+ , тропониновый комплекс претерпевает конформационные изменения, вызывающие смещение тропомиозина с участков связывания миозина на F-актин, что позволяет связываться миозину толстых филаментов.
В результате связывания Ca2+ , тропониновый комплекс претерпевает конформационные изменения, вызывающие смещение тропомиозина с участков связывания миозина на F-актин, что позволяет связываться миозину толстых филаментов.
Цикл поперечного моста, событие, происходящее во время сопряжения возбуждения и сокращения, относится к механизму, с помощью которого толстые и тонкие нити скользят относительно друг друга, вызывая мышечное сокращение. В начале цикла, когда миозин прочно связан с актином, аденозинтрифосфат (АТФ) не связан с миозином, состояние, известное как ригидность; это преходящее состояние при сокращении мышц, тогда как при отсутствии АТФ, например, при смерти, это состояние является постоянным и называется трупным окоченением. Затем АТФ связывается с головкой миозина, вызывая конформационные изменения миозина, которые снижают его сродство к актину. Следовательно, миозин отделяется от актина, и миозиновая головка наклоняется к концу саркомера. АТФ, связанный с миозином, гидролизуется до аденозиндифосфата (АДФ) и одной молекулы неорганического фосфата, которые остаются связанными с миозином. Во взведенном положении миозин затем связывается с новым участком на актине, создавая силовой удар, который тянет актиновые филаменты. Каждый цикл перекрестного мостика приводит к тому, что головка миозина продвигается вверх по актиновому филаменту при условии, что Са2+ остается связанным с тропонином С. отсутствие АТФ.[4]
Во взведенном положении миозин затем связывается с новым участком на актине, создавая силовой удар, который тянет актиновые филаменты. Каждый цикл перекрестного мостика приводит к тому, что головка миозина продвигается вверх по актиновому филаменту при условии, что Са2+ остается связанным с тропонином С. отсутствие АТФ.[4]
После сокращения происходит расслабление мышц, когда Са2+ повторно накапливается в саркоплазматическом ретикулуме с помощью активного насоса Са2+-АТФазы (SERCA) на мембране саркоплазматического ретикулума. Этот насос переносит внутриклеточный Са2+ в саркоплазматический ретикулум, который поддерживает низкий уровень внутриклеточного Са2+, когда мышца расслаблена. В саркоплазматическом ретикулуме находится белок, связывающий Са2+, называемый кальсеквестрин, который служит для снижения концентрации свободного Са2+, чтобы уменьшить объем работы, необходимой насосу SERCA. Когда внутриклеточная концентрация Са2+ снижается, Са2+ диссоциирует от тропонина С, позволяя тропомиозину возобновить блокирование участков связывания миозина на F-актине.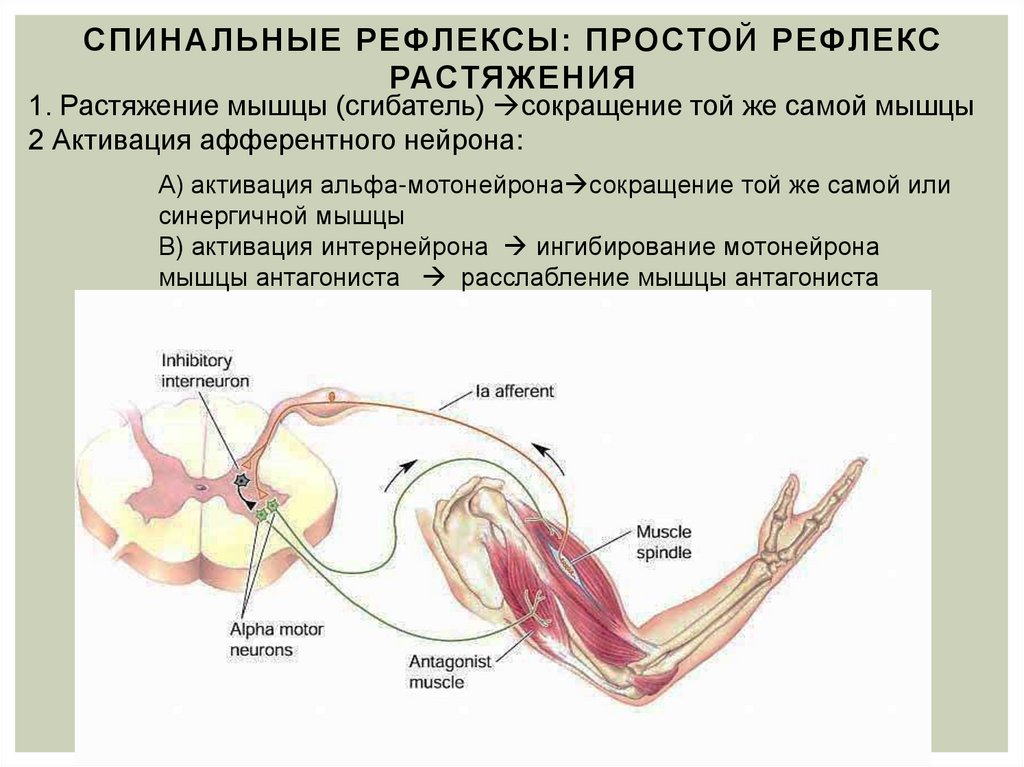 [5]
[5]
События сопряжения возбуждения и сокращения всегда следуют друг за другом и демонстрируют временную зависимость. Другими словами, потенциал действия мышечного волокна всегда предшествует увеличению внутриклеточного Са2+, что всегда предшествует мышечному сокращению. Один единственный потенциал действия, приводящий к увеличению внутриклеточного Ca2+ за счет высвобождения саркоплазматического ретикулума, вызывает одиночное мышечное сокращение, известное как подергивание. Поскольку продолжительность потенциала действия короче продолжительности сокращения, мышечное волокно может снова активироваться до того, как произойдет мышечное расслабление. Если уже активное мышечное волокно снова стимулируется, у саркоплазматического ретикулума недостаточно времени для повторного накопления Ca2+. Следовательно, внутриклеточный Ca2+ остается высоким, а сила второго стимула становится аддитивным эффектом к остальной части первого стимула, что приводит к дополнительной силе. Это явление длительного сокращения называется тетанией.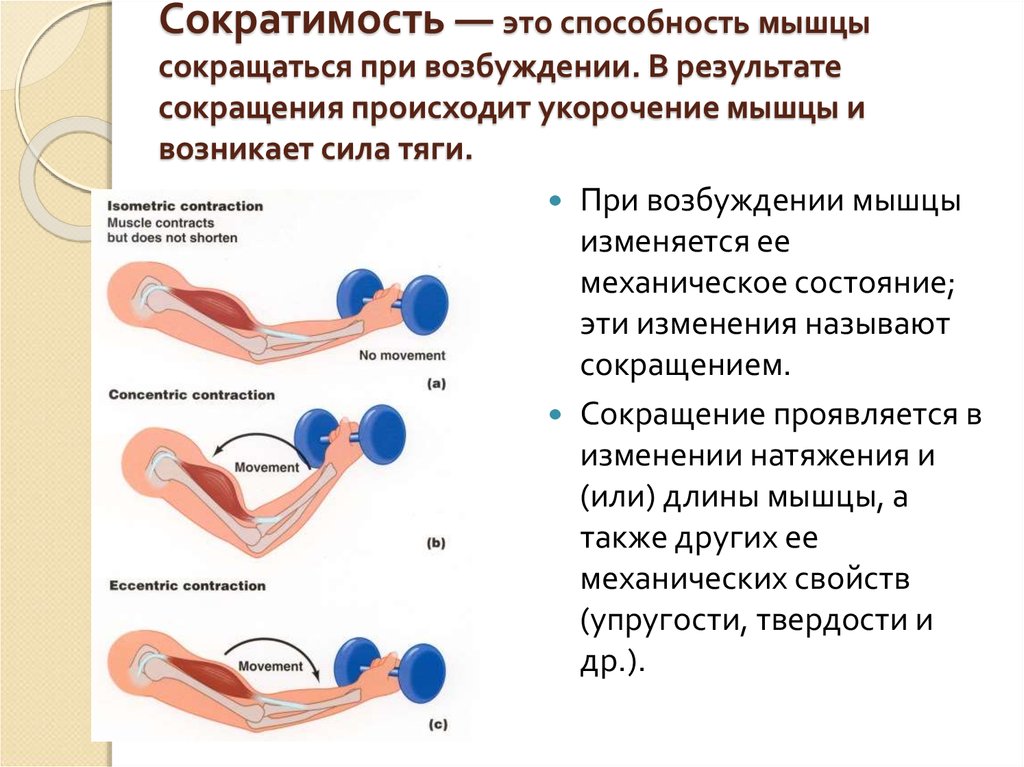
Соотношение длина-напряжение в мышцах иллюстрирует напряжения или силы, возникающие в цикле поперечного моста в результате изменений длины мышечных волокон. Напряжение определяется путем изменения длины покоя мышцы, которая уже подверглась изометрическому сокращению. Таким образом, эта длина в состоянии покоя, также известная как предварительная нагрузка, является результатом пассивного предварительного сокращения изометрического сокращения. Пассивное напряжение относится к напряжению, которое возникает просто из-за увеличения длины мышцы. По мере увеличения преднагрузки и удлинения мышцы ее напряжение еще больше возрастает. Пассивное натяжение можно рассматривать как натяжение эластичной резиновой ленты по мере ее дальнейшего растяжения. Активное натяжение — это натяжение, возникающее в результате цикла поперечных мостов, и оно пропорционально фактическому количеству поперечных мостов. Это напряжение является самым высоким, когда существует оптимальное перекрытие между миозином и актином, что приводит к максимальному количеству поперечных мостиков.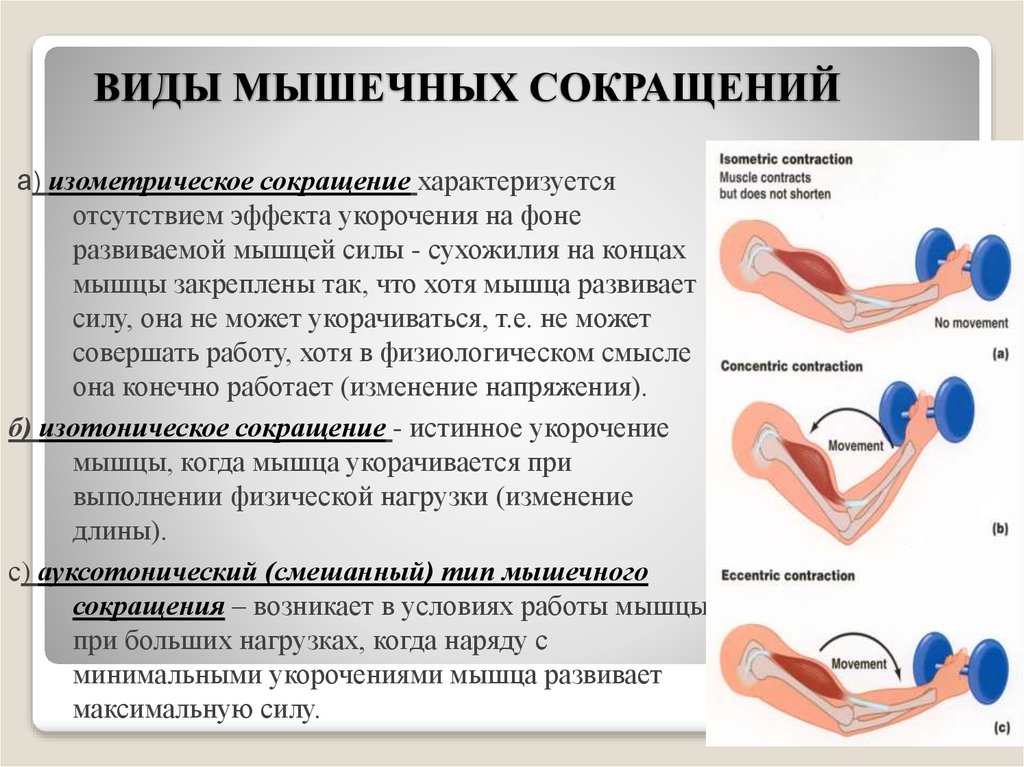 Когда длина мышцы уменьшается, происходит скученность нитей, что снижает напряжение. Точно так же, когда длина мышцы увеличивается, активное напряжение уменьшается из-за меньшего перекрытия между миозином и актином и, соответственно, меньшего количества поперечных мостиков. Общее напряжение представляет собой напряжение, возникающее в результате сокращения мышц при различных предварительных нагрузках, и равно сумме активного напряжения и пассивного напряжения.[6]
Когда длина мышцы уменьшается, происходит скученность нитей, что снижает напряжение. Точно так же, когда длина мышцы увеличивается, активное напряжение уменьшается из-за меньшего перекрытия между миозином и актином и, соответственно, меньшего количества поперечных мостиков. Общее напряжение представляет собой напряжение, возникающее в результате сокращения мышц при различных предварительных нагрузках, и равно сумме активного напряжения и пассивного напряжения.[6]
Соотношение сила-скорость относится к скорости сокращения мышц как функции постнагрузки, которая представляет собой силу, против которой мышца сокращается. В этом отношении постнагрузка является фиксированной переменной, в отличие от отношения длины к напряжению, когда фиксированной переменной была длина мышцы. По мере увеличения постнагрузки скорость укорочения уменьшается. Максимальная скорость достигается при нулевой постнагрузке мышцы.[7]
Концентрическое сокращение означает, что сила сокращения превышает силу сопротивления, что приводит к укорочению мышцы и сближению начала и прикрепления мышцы. Эксцентрическое сокращение возникает, когда сила сокращения меньше силы сопротивления. Другими словами, сила сопротивления больше, чем сила сокращения, что приводит к удлинению мышцы и увеличению расстояния между ее началом и местом прикрепления.
Эксцентрическое сокращение возникает, когда сила сокращения меньше силы сопротивления. Другими словами, сила сопротивления больше, чем сила сокращения, что приводит к удлинению мышцы и увеличению расстояния между ее началом и местом прикрепления.
Патофизиология
Злокачественная гипертермия представляет собой опасное для жизни состояние, возникающее преимущественно у лиц с генетической предрасположенностью с мутацией рианодинового рецептора саркоплазматического ретикулума. Когда эти люди подвергаются воздействию летучих анестетиков или миорелаксанта сукцинилхолина, происходит массивное высвобождение внутриклеточного Ca2+ из рианодиновых рецепторов и недостаточная секвестрация Ca2+ насосом SERCA. Этот механизм приводит к сокращению мышц, рабдомиолизу, тяжелой гипертермии и, возможно, смерти. Единственным средством лечения злокачественной гипертермии является дантролен, который связывается с рианодиновым рецептором, чтобы предотвратить дальнейшее высвобождение Ca2+.[8]
Миастения гравис — это аутоиммунное заболевание, поражающее нервно-мышечное соединение. Он характеризуется утомляемой слабостью скелетных мышц, которая усиливается при повторяющихся движениях и улучшается в покое. Чаще всего миастения первоначально включает слабость глазных мышц с возможным прогрессированием на мышцы конечностей. У большинства пациентов с этим заболеванием выявляются аутоантитела к никотиновым АХ-рецепторам нервно-мышечного синапса, что вызывает эндоцитоз и деградацию рецепторов. Без связывания АХ с рецепторами потенциалы действия не могут распространяться вниз по мышечному волокну, и, следовательно, возникает мышечная слабость. Ингибиторы ацетилхолинэстеразы предотвращают распад ацетилхолина и используются для усиления нервно-мышечной передачи при лечении миастении [9].]
Он характеризуется утомляемой слабостью скелетных мышц, которая усиливается при повторяющихся движениях и улучшается в покое. Чаще всего миастения первоначально включает слабость глазных мышц с возможным прогрессированием на мышцы конечностей. У большинства пациентов с этим заболеванием выявляются аутоантитела к никотиновым АХ-рецепторам нервно-мышечного синапса, что вызывает эндоцитоз и деградацию рецепторов. Без связывания АХ с рецепторами потенциалы действия не могут распространяться вниз по мышечному волокну, и, следовательно, возникает мышечная слабость. Ингибиторы ацетилхолинэстеразы предотвращают распад ацетилхолина и используются для усиления нервно-мышечной передачи при лечении миастении [9].]
Ботулинический токсин – это агент, который изменяет нервно-мышечную функцию. Этот токсин, продуцируемый C. botulinum , препятствует высвобождению АХ из пресинаптической мембраны мотонейрона. Следовательно, скелетные мышцы не могут сокращаться, что приводит к вялому параличу. [10]
[10]
Судороги скелетных мышц возникают из-за внезапных и непроизвольных сокращений мышц, которые длятся от секунд до минут и вызывают боль. Хотя они могут быть связаны с заболеваниями, чаще всего мышечные судороги возникают при отсутствии какой-либо явной патологии. Они чаще всего проявляются у пациентов пожилого возраста, беременных или активно занимающихся физическими упражнениями. Поскольку судороги являются результатом мышечных сокращений, немедленное облегчение боли может быть обеспечено путем растяжения пораженной мышцы.[11]
Клиническое значение
Оценка мышечной силы и мышечного сокращения является стандартной процедурой, включаемой в физикальное обследование пациента. Шкала ручного мышечного тестирования Совета медицинских исследований является наиболее часто используемой системой оценки мышечной силы, где баллы от 0 до 5 присваиваются в зависимости от способностей пациента. Оценка 0 означает отсутствие активации мышц. Оценка 1 означает, что имеется лишь небольшая сократимость мышцы.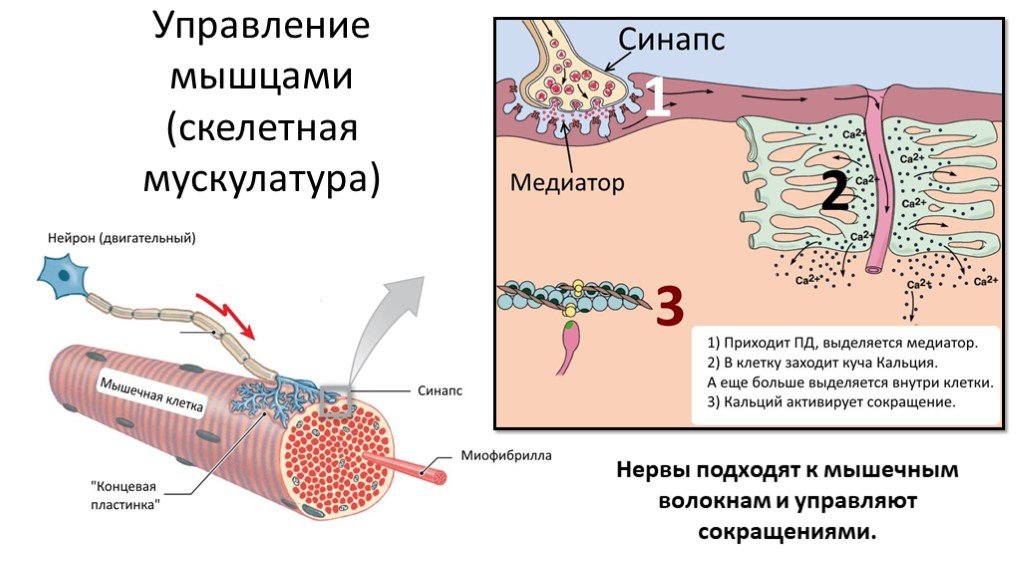 2 балла — это когда активация мышц тестируется в отсутствие гравитации. 3 относится к активации мышц против силы тяжести, но не с сопротивлением. 4 — активация мышц против силы тяжести и некоторого сопротивления, а 5 — активация мышц против гравитации и полного сопротивления. Основная цель проверки мышечной силы при физикальном обследовании состоит в том, чтобы оценить и установить дифференциальный диагноз, когда пациент предъявляет жалобы на слабость, часто на фоне неврологического заболевания. [12]
2 балла — это когда активация мышц тестируется в отсутствие гравитации. 3 относится к активации мышц против силы тяжести, но не с сопротивлением. 4 — активация мышц против силы тяжести и некоторого сопротивления, а 5 — активация мышц против гравитации и полного сопротивления. Основная цель проверки мышечной силы при физикальном обследовании состоит в том, чтобы оценить и установить дифференциальный диагноз, когда пациент предъявляет жалобы на слабость, часто на фоне неврологического заболевания. [12]
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.
Комментарий к этой статье.
Ссылки
- 1.
Сквайр Дж. Специальный выпуск: Актин-миозиновое взаимодействие в мышцах: история и обзор. Int J Mol Sci. 14 ноября 2019 г. 20 (22) [Бесплатная статья PMC: PMC6887992] [PubMed: 31739584]
- 2.
Оцуки И., Моримото С. Тропонин: регуляторная функция и нарушения.
Biochem Biophys Res Commun. 2008 25 апреля; 369(1):62-73. [PubMed: 18154728]- 3.
Сантулли Г., Льюис Д.Р., Маркс А.Р. Физиология и патофизиология сопряжения возбуждения и сокращения: функциональная роль рианодинового рецептора. J Muscle Res Cell Motil. 2017 фев;38(1):37-45. [Бесплатная статья PMC: PMC5813681] [PubMed: 28653141]
- 4.
Fitts RH. Цикл поперечного моста и усталость скелетных мышц. J Appl Physiol (1985). 2008 г., февраль; 104 (2): 551-8. [PubMed: 18162480]
- 5.
Paolini C, Quarta M, Nori A, Boncompagni S, Canato M, Volpe P, Allen PD, Reggiani C, Protasi F. Реорганизованные запасы и нарушение обработки кальция в скелетных мышцах мышей, лишенных кальсеквестрина-1. Дж. Физиол. 01 сентября 2007 г .: 583 (часть 2): 767–84. [Бесплатная статья PMC: PMC2277031] [PubMed: 17627988]
- 6.
Lieber RL, Ward SR. Конструкция скелетных мышц для удовлетворения функциональных требований.
Philos Trans R Soc Lond B Biol Sci. 2011 27 мая; 366 (1570): 1466-76. [Бесплатная статья PMC: PMC3130443] [PubMed: 21502118]- 7.
Джозефсон Р.К., Эдман К.А. Изменение максимальной скорости укорочения мышечных волокон лягушки в начале тетанического сокращения и во время расслабления. Дж. Физиол. 1998 01 марта; 507 (часть 2) (часть 2): 511-25. [Бесплатная статья PMC: PMC2230787] [PubMed: 9518709]
- 8.
Розенберг Х., Поллок Н., Шиманн А., Балгер Т., Стоуэлл К. Злокачественная гипертермия: обзор. Orphanet J Rare Dis. 2015 04 авг;10:93. [Бесплатная статья PMC: PMC4524368] [PubMed: 26238698]
- 9.
Koneczny I, Herbst R. Myasthenia Gravis: Патогенные эффекты аутоантител на нервно-мышечную архитектуру. Клетки. 2019 Jul 02;8(7) [Бесплатная статья PMC: PMC6678492] [PubMed: 31269763]
- 10.
Дресслер Д., Сабери Ф.А., Барбоза Э.Р. Ботулинический токсин: механизмы действия. Арк Нейропсиквиатр.
2005 март; 63(1):180-5. [PubMed: 15830090]- 11.
Бордони Б., Сугумар К., Варакалло М. StatPearls [Интернет]. Издательство StatPearls; Остров сокровищ (Флорида): 4 сентября 2022 г. Мышечные спазмы. [В паблике: 29763070]
- 12.
Накви У, Шерман А.Л. StatPearls [Интернет]. Издательство StatPearls; Остров сокровищ (Флорида): 29 августа 2022 г. Оценка мышечной силы. [PubMed: 28613779]
 Biochem Biophys Res Commun. 2008 25 апреля; 369(1):62-73. [PubMed: 18154728]
Biochem Biophys Res Commun. 2008 25 апреля; 369(1):62-73. [PubMed: 18154728] Philos Trans R Soc Lond B Biol Sci. 2011 27 мая; 366 (1570): 1466-76. [Бесплатная статья PMC: PMC3130443] [PubMed: 21502118]
Philos Trans R Soc Lond B Biol Sci. 2011 27 мая; 366 (1570): 1466-76. [Бесплатная статья PMC: PMC3130443] [PubMed: 21502118] 2005 март; 63(1):180-5. [PubMed: 15830090]
2005 март; 63(1):180-5. [PubMed: 15830090]Физиология, сокращение скелетных мышц — StatPearls
Введение
Как органы, содержащие клетки, способные сокращаться, мышцы могут генерировать силу и движение. Скелетные мышцы работают вместе с костями скелета, создавая движения тела. Кроме того, он также связан с мышцами диафрагмы, пищевода и глаз. Таким образом, скелетные мышцы служат множеству целей, включая движение тела, дыхание и глотание. В отличие от гладких мышц и сердечной мышцы, скелетные мышцы сокращаются в основном в ответ на произвольные стимулы.
Клеточный
Скелетная мышца состоит из клеток, которые в совокупности называются мышечными волокнами.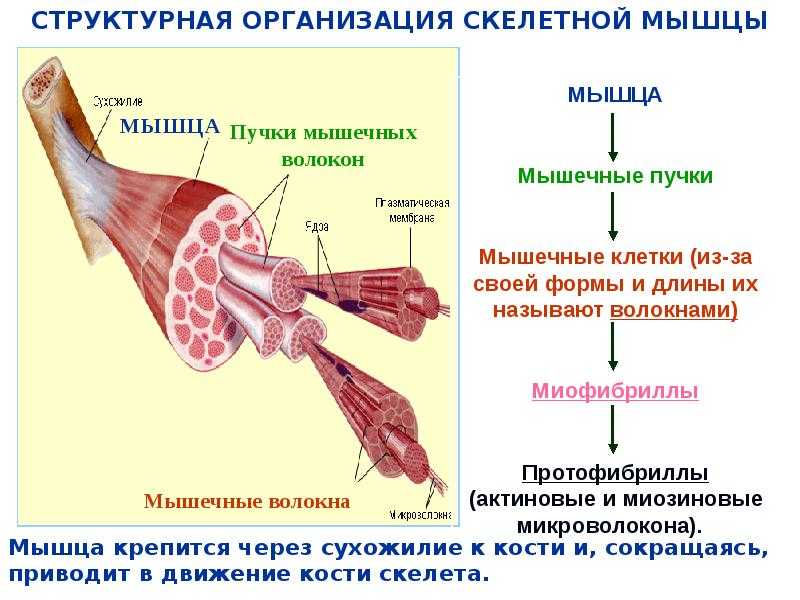 Каждое мышечное волокно многоядерное, его ядра расположены по периферии волокна. Каждое мышечное волокно далее подразделяется на миофибриллы, которые являются основными единицами мышечного волокна. Эти миофибриллы окружены мышечной клеточной мембраной (сарколеммой), которая образует внутри миофибриллы глубокие впячивания, называемые поперечными трубочками (Т-трубочками). Каждая миофибрилла содержит сократительные белки, описываемые как толстые и тонкие филаменты, которые расположены продольно в единицы, называемые саркомерами.
Каждое мышечное волокно многоядерное, его ядра расположены по периферии волокна. Каждое мышечное волокно далее подразделяется на миофибриллы, которые являются основными единицами мышечного волокна. Эти миофибриллы окружены мышечной клеточной мембраной (сарколеммой), которая образует внутри миофибриллы глубокие впячивания, называемые поперечными трубочками (Т-трубочками). Каждая миофибрилла содержит сократительные белки, описываемые как толстые и тонкие филаменты, которые расположены продольно в единицы, называемые саркомерами.
Основной единицей толстого филамента является большой белок миозин, который образован двумя парами легких цепей и одной парой тяжелых цепей. Две тяжелые цепи миозина закручиваются друг вокруг друга, образуя спиральный хвост миозина, тогда как легкие цепи взаимодействуют с тяжелыми цепями, образуя две головки миозина на другом конце. На головках находится важный участок связывания, который облегчает взаимодействие миозина с актином, белком, принадлежащим тонкому филаменту.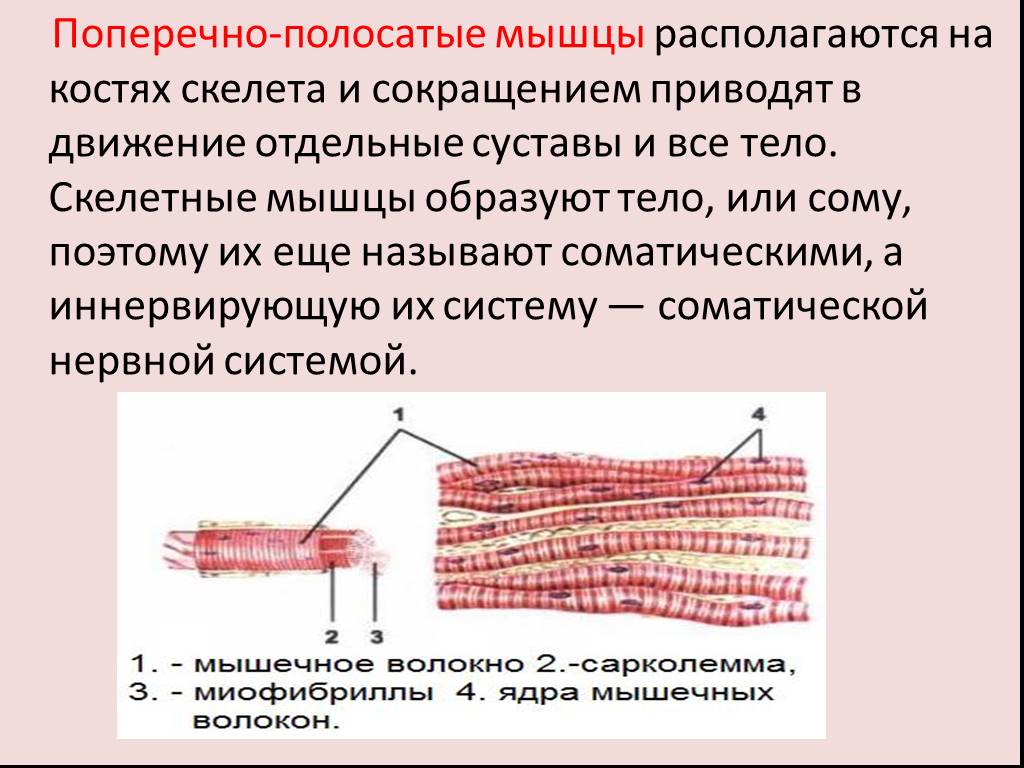 [1]
[1]
Другой сократительной нитью в миофибриллах является тонкая нить, в основном состоящая из трех белков: актина, тропомиозина и тропонина. Мономерная глобулярная форма актина, называемая G-актином, полимеризуется в две нити, которые скручиваются и переплетаются друг с другом, образуя нитевидный актин, называемый F-актином. По всей длине F-актина находятся сайты связывания миозина, которые скрыты нитчатым белком тропомиозином. Функция тропомиозина заключается в предотвращении взаимодействия актина и миозина, когда мышца находится в состоянии покоя, что, следовательно, предотвращает сокращение мышцы. Тропонин представляет собой комплекс из трех белков, расположенных вдоль тропомиозиновых филаментов. Первый белок, тропонин Т, способствует связыванию тропонина с тропомиозином. Тропонин I служит той же цели, что и тропомиозин, останавливая актин-миозиновое взаимодействие, блокируя сайты связывания миозина. Наконец, тропонин С связывает кальций, чтобы инициировать мышечное сокращение.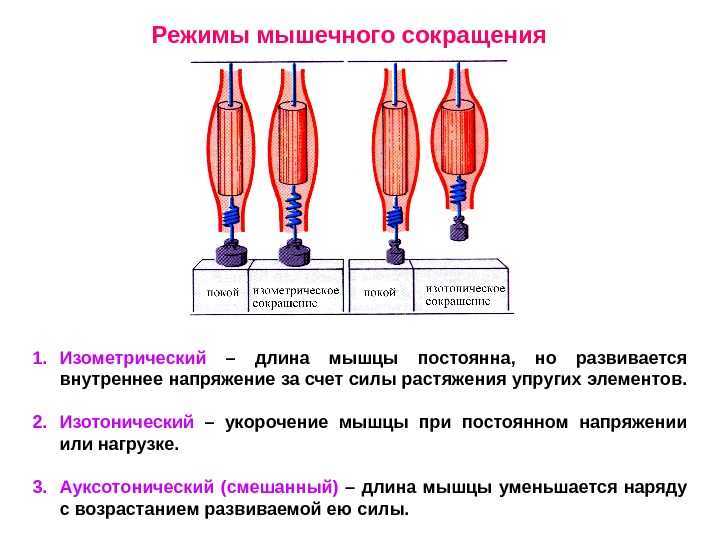 [2]
[2]
Как упоминалось ранее, толстые и тонкие нити миофибрилл организованы в единицы, называемые саркомерами. Саркомер является основной сократительной единицей миофибриллы. Z-линии разделяют каждый саркомер. Полосы А, расположенные в центре каждого саркомера, содержат толстые филаменты, которые могут перекрываться с тонкими филаментами. Полоса А далее делится на зону Н, которая не содержит тонких нитей. Выступающая линия М делит пополам зону Н и служит для соединения средних частей толстых нитей. По обеим сторонам полосы A расположены полосы I, которые содержат как тонкие нити, так и линию Z, которая проходит по середине каждой полосы I.
Механизм
Нервы, отвечающие за иннервацию мышечных волокон, называются мотонейронами. Отдельный мотонейрон и мышечные волокна, которые он иннервирует, в совокупности называются двигательной единицей. Количество мышечных волокон в двигательной единице предсказуемо зависит от функции мышцы. Например, двигательные единицы, ответственные за мимические мышцы, включают в себя значительно меньше мышечных волокон, чем двигательные единицы, отвечающие за мышцы, участвующие в таких действиях, как плавание.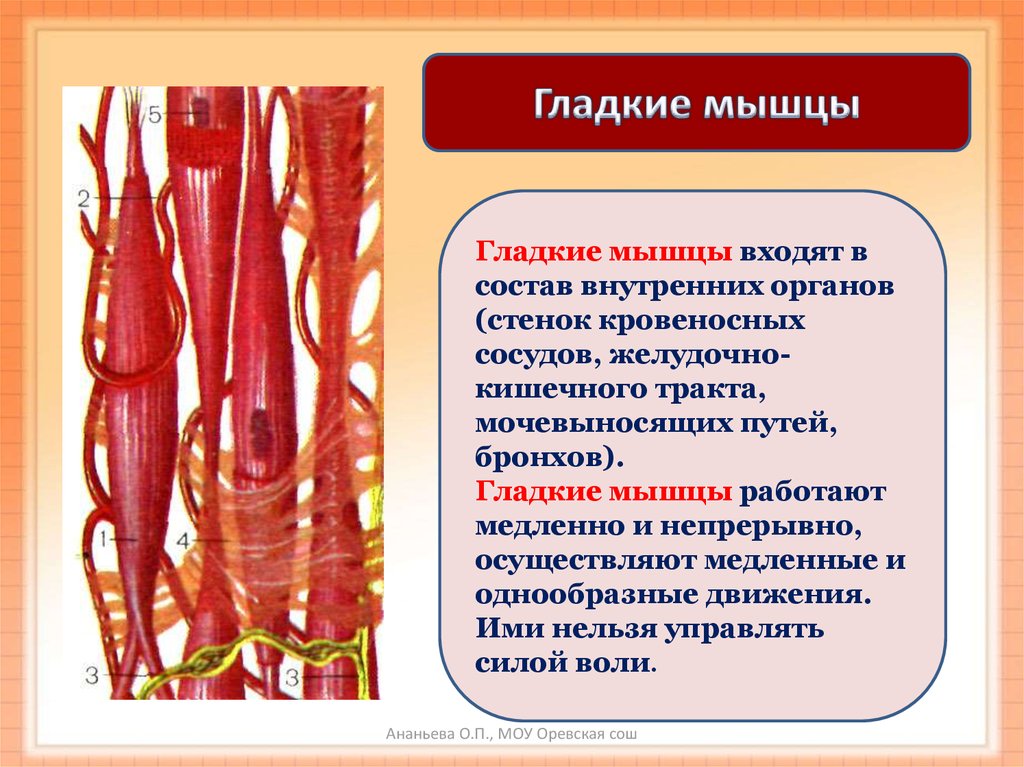
Сокращение скелетных мышц начинается сначала в нервно-мышечном соединении, которое является синапсом между мотонейроном и мышечным волокном. Распространение потенциалов действия на мотонейрон и последующая деполяризация приводят к открытию потенциалзависимых кальциевых (Са2+) каналов пресинаптической мембраны. Поток Са2+ внутрь вызывает высвобождение ацетилхолина (АХ) в нервно-мышечном соединении, который диффундирует к постсинаптической мембране мышечного волокна. Постсинаптическая мембрана мышечного волокна также известна как двигательная замыкательная пластинка. АХ связывается с никотиновыми рецепторами, расположенными на двигательной концевой пластинке, деполяризуя ее, что инициирует потенциалы действия в мышечном волокне.
Связь возбуждения и сокращения относится к механизму, который преобразует упомянутые выше потенциалы действия в мышечных волокнах в сокращение мышечных волокон. Потенциалы действия на мембране мышечных клеток, окружающих миофибриллы, проходят в Т-трубочки, которые отвечают за распространение потенциала действия с поверхности внутрь мышечного волокна.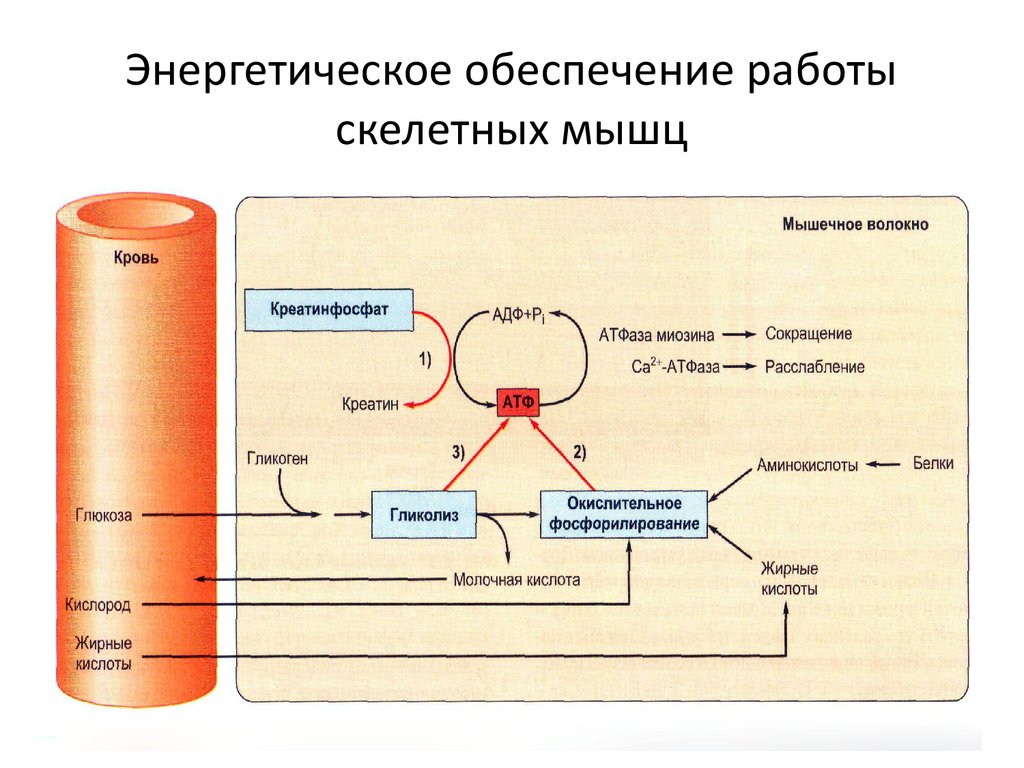 Т-трубочки содержат дигидропиридиновые рецепторы, которые прилегают к терминальным цистернам саркоплазматического ретикулума мышечного волокна. Когда Т-трубочки деполяризуются, их дигидропиридиновые рецепторы претерпевают конформационные изменения, которые механически взаимодействуют с рианодиновыми рецепторами на саркоплазматическом ретикулуме. Это взаимодействие открывает рианодиновые рецепторы, вызывая высвобождение Ca2+ из саркоплазматического ретикулума. В результате повышенный внутриклеточный Са2+ прикрепляется к тропонину С тропонинового комплекса на тонких филаментах. Взаимодействие между Ca2+ и тропонином C проявляет кооперативность, что означает, что каждый Ca2+, который связывает тропонин C, увеличивает сродство связывания тропонина C со следующей молекулой Ca2+, всего до четырех ионов Ca2+ на тропонин C. В результате связывания Ca2+ , тропониновый комплекс претерпевает конформационные изменения, вызывающие смещение тропомиозина с участков связывания миозина на F-актин, что позволяет связываться миозину толстых филаментов.
Т-трубочки содержат дигидропиридиновые рецепторы, которые прилегают к терминальным цистернам саркоплазматического ретикулума мышечного волокна. Когда Т-трубочки деполяризуются, их дигидропиридиновые рецепторы претерпевают конформационные изменения, которые механически взаимодействуют с рианодиновыми рецепторами на саркоплазматическом ретикулуме. Это взаимодействие открывает рианодиновые рецепторы, вызывая высвобождение Ca2+ из саркоплазматического ретикулума. В результате повышенный внутриклеточный Са2+ прикрепляется к тропонину С тропонинового комплекса на тонких филаментах. Взаимодействие между Ca2+ и тропонином C проявляет кооперативность, что означает, что каждый Ca2+, который связывает тропонин C, увеличивает сродство связывания тропонина C со следующей молекулой Ca2+, всего до четырех ионов Ca2+ на тропонин C. В результате связывания Ca2+ , тропониновый комплекс претерпевает конформационные изменения, вызывающие смещение тропомиозина с участков связывания миозина на F-актин, что позволяет связываться миозину толстых филаментов.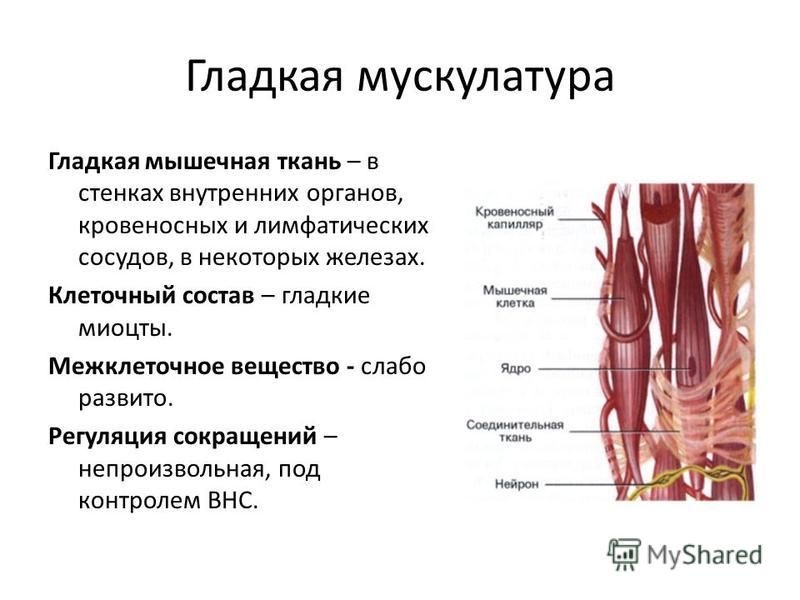
Цикл поперечного моста, событие, происходящее во время сопряжения возбуждения и сокращения, относится к механизму, с помощью которого толстые и тонкие нити скользят относительно друг друга, вызывая мышечное сокращение. В начале цикла, когда миозин прочно связан с актином, аденозинтрифосфат (АТФ) не связан с миозином, состояние, известное как ригидность; это преходящее состояние при сокращении мышц, тогда как при отсутствии АТФ, например, при смерти, это состояние является постоянным и называется трупным окоченением. Затем АТФ связывается с головкой миозина, вызывая конформационные изменения миозина, которые снижают его сродство к актину. Следовательно, миозин отделяется от актина, и миозиновая головка наклоняется к концу саркомера. АТФ, связанный с миозином, гидролизуется до аденозиндифосфата (АДФ) и одной молекулы неорганического фосфата, которые остаются связанными с миозином. Во взведенном положении миозин затем связывается с новым участком на актине, создавая силовой удар, который тянет актиновые филаменты. Каждый цикл перекрестного мостика приводит к тому, что головка миозина продвигается вверх по актиновому филаменту при условии, что Са2+ остается связанным с тропонином С. отсутствие АТФ.[4]
Каждый цикл перекрестного мостика приводит к тому, что головка миозина продвигается вверх по актиновому филаменту при условии, что Са2+ остается связанным с тропонином С. отсутствие АТФ.[4]
После сокращения происходит расслабление мышц, когда Са2+ повторно накапливается в саркоплазматическом ретикулуме с помощью активного насоса Са2+-АТФазы (SERCA) на мембране саркоплазматического ретикулума. Этот насос переносит внутриклеточный Са2+ в саркоплазматический ретикулум, который поддерживает низкий уровень внутриклеточного Са2+, когда мышца расслаблена. В саркоплазматическом ретикулуме находится белок, связывающий Са2+, называемый кальсеквестрин, который служит для снижения концентрации свободного Са2+, чтобы уменьшить объем работы, необходимой насосу SERCA. Когда внутриклеточная концентрация Са2+ снижается, Са2+ диссоциирует от тропонина С, позволяя тропомиозину возобновить блокирование участков связывания миозина на F-актине.[5]
События сопряжения возбуждения и сокращения всегда следуют друг за другом и демонстрируют временную зависимость. Другими словами, потенциал действия мышечного волокна всегда предшествует увеличению внутриклеточного Са2+, что всегда предшествует мышечному сокращению. Один единственный потенциал действия, приводящий к увеличению внутриклеточного Ca2+ за счет высвобождения саркоплазматического ретикулума, вызывает одиночное мышечное сокращение, известное как подергивание. Поскольку продолжительность потенциала действия короче продолжительности сокращения, мышечное волокно может снова активироваться до того, как произойдет мышечное расслабление. Если уже активное мышечное волокно снова стимулируется, у саркоплазматического ретикулума недостаточно времени для повторного накопления Ca2+. Следовательно, внутриклеточный Ca2+ остается высоким, а сила второго стимула становится аддитивным эффектом к остальной части первого стимула, что приводит к дополнительной силе. Это явление длительного сокращения называется тетанией.
Другими словами, потенциал действия мышечного волокна всегда предшествует увеличению внутриклеточного Са2+, что всегда предшествует мышечному сокращению. Один единственный потенциал действия, приводящий к увеличению внутриклеточного Ca2+ за счет высвобождения саркоплазматического ретикулума, вызывает одиночное мышечное сокращение, известное как подергивание. Поскольку продолжительность потенциала действия короче продолжительности сокращения, мышечное волокно может снова активироваться до того, как произойдет мышечное расслабление. Если уже активное мышечное волокно снова стимулируется, у саркоплазматического ретикулума недостаточно времени для повторного накопления Ca2+. Следовательно, внутриклеточный Ca2+ остается высоким, а сила второго стимула становится аддитивным эффектом к остальной части первого стимула, что приводит к дополнительной силе. Это явление длительного сокращения называется тетанией.
Соотношение длина-напряжение в мышцах иллюстрирует напряжения или силы, возникающие в цикле поперечного моста в результате изменений длины мышечных волокон. Напряжение определяется путем изменения длины покоя мышцы, которая уже подверглась изометрическому сокращению. Таким образом, эта длина в состоянии покоя, также известная как предварительная нагрузка, является результатом пассивного предварительного сокращения изометрического сокращения. Пассивное напряжение относится к напряжению, которое возникает просто из-за увеличения длины мышцы. По мере увеличения преднагрузки и удлинения мышцы ее напряжение еще больше возрастает. Пассивное натяжение можно рассматривать как натяжение эластичной резиновой ленты по мере ее дальнейшего растяжения. Активное натяжение — это натяжение, возникающее в результате цикла поперечных мостов, и оно пропорционально фактическому количеству поперечных мостов. Это напряжение является самым высоким, когда существует оптимальное перекрытие между миозином и актином, что приводит к максимальному количеству поперечных мостиков. Когда длина мышцы уменьшается, происходит скученность нитей, что снижает напряжение. Точно так же, когда длина мышцы увеличивается, активное напряжение уменьшается из-за меньшего перекрытия между миозином и актином и, соответственно, меньшего количества поперечных мостиков.
Напряжение определяется путем изменения длины покоя мышцы, которая уже подверглась изометрическому сокращению. Таким образом, эта длина в состоянии покоя, также известная как предварительная нагрузка, является результатом пассивного предварительного сокращения изометрического сокращения. Пассивное напряжение относится к напряжению, которое возникает просто из-за увеличения длины мышцы. По мере увеличения преднагрузки и удлинения мышцы ее напряжение еще больше возрастает. Пассивное натяжение можно рассматривать как натяжение эластичной резиновой ленты по мере ее дальнейшего растяжения. Активное натяжение — это натяжение, возникающее в результате цикла поперечных мостов, и оно пропорционально фактическому количеству поперечных мостов. Это напряжение является самым высоким, когда существует оптимальное перекрытие между миозином и актином, что приводит к максимальному количеству поперечных мостиков. Когда длина мышцы уменьшается, происходит скученность нитей, что снижает напряжение. Точно так же, когда длина мышцы увеличивается, активное напряжение уменьшается из-за меньшего перекрытия между миозином и актином и, соответственно, меньшего количества поперечных мостиков.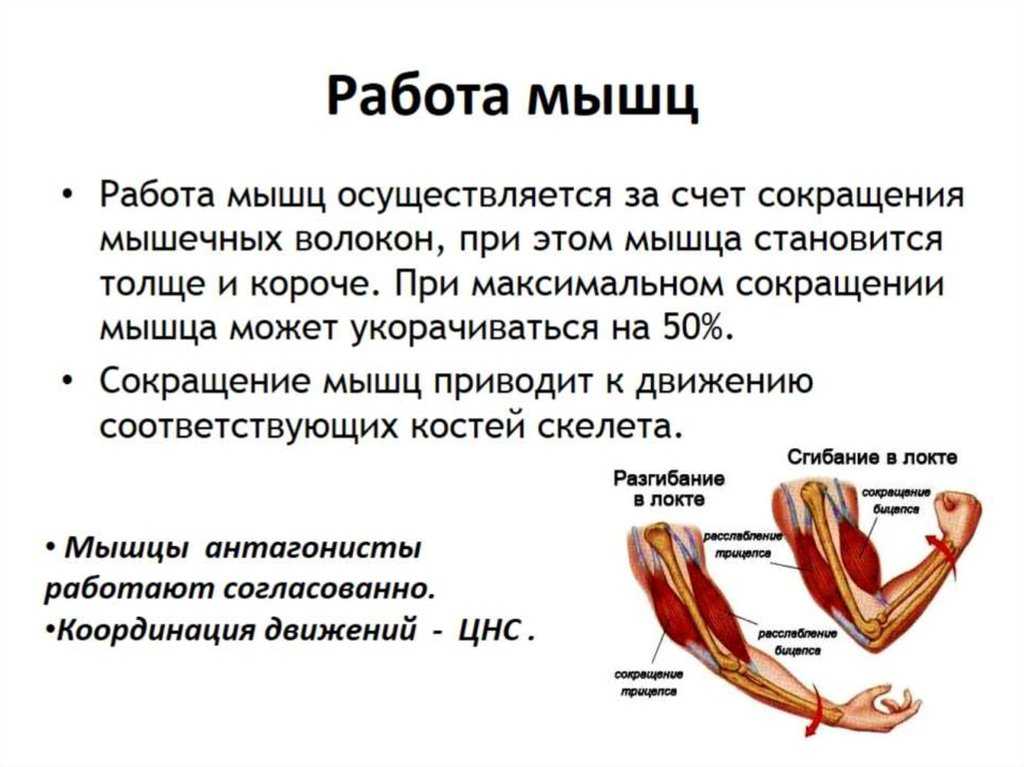 Общее напряжение представляет собой напряжение, возникающее в результате сокращения мышц при различных предварительных нагрузках, и равно сумме активного напряжения и пассивного напряжения.[6]
Общее напряжение представляет собой напряжение, возникающее в результате сокращения мышц при различных предварительных нагрузках, и равно сумме активного напряжения и пассивного напряжения.[6]
Соотношение сила-скорость относится к скорости сокращения мышц как функции постнагрузки, которая представляет собой силу, против которой мышца сокращается. В этом отношении постнагрузка является фиксированной переменной, в отличие от отношения длины к напряжению, когда фиксированной переменной была длина мышцы. По мере увеличения постнагрузки скорость укорочения уменьшается. Максимальная скорость достигается при нулевой постнагрузке мышцы.[7]
Концентрическое сокращение означает, что сила сокращения превышает силу сопротивления, что приводит к укорочению мышцы и сближению начала и прикрепления мышцы. Эксцентрическое сокращение возникает, когда сила сокращения меньше силы сопротивления. Другими словами, сила сопротивления больше, чем сила сокращения, что приводит к удлинению мышцы и увеличению расстояния между ее началом и местом прикрепления.
Патофизиология
Злокачественная гипертермия представляет собой опасное для жизни состояние, возникающее преимущественно у лиц с генетической предрасположенностью с мутацией рианодинового рецептора саркоплазматического ретикулума. Когда эти люди подвергаются воздействию летучих анестетиков или миорелаксанта сукцинилхолина, происходит массивное высвобождение внутриклеточного Ca2+ из рианодиновых рецепторов и недостаточная секвестрация Ca2+ насосом SERCA. Этот механизм приводит к сокращению мышц, рабдомиолизу, тяжелой гипертермии и, возможно, смерти. Единственным средством лечения злокачественной гипертермии является дантролен, который связывается с рианодиновым рецептором, чтобы предотвратить дальнейшее высвобождение Ca2+.[8]
Миастения гравис — это аутоиммунное заболевание, поражающее нервно-мышечное соединение. Он характеризуется утомляемой слабостью скелетных мышц, которая усиливается при повторяющихся движениях и улучшается в покое. Чаще всего миастения первоначально включает слабость глазных мышц с возможным прогрессированием на мышцы конечностей. У большинства пациентов с этим заболеванием выявляются аутоантитела к никотиновым АХ-рецепторам нервно-мышечного синапса, что вызывает эндоцитоз и деградацию рецепторов. Без связывания АХ с рецепторами потенциалы действия не могут распространяться вниз по мышечному волокну, и, следовательно, возникает мышечная слабость. Ингибиторы ацетилхолинэстеразы предотвращают распад ацетилхолина и используются для усиления нервно-мышечной передачи при лечении миастении [9].]
У большинства пациентов с этим заболеванием выявляются аутоантитела к никотиновым АХ-рецепторам нервно-мышечного синапса, что вызывает эндоцитоз и деградацию рецепторов. Без связывания АХ с рецепторами потенциалы действия не могут распространяться вниз по мышечному волокну, и, следовательно, возникает мышечная слабость. Ингибиторы ацетилхолинэстеразы предотвращают распад ацетилхолина и используются для усиления нервно-мышечной передачи при лечении миастении [9].]
Ботулинический токсин – это агент, который изменяет нервно-мышечную функцию. Этот токсин, продуцируемый C. botulinum , препятствует высвобождению АХ из пресинаптической мембраны мотонейрона. Следовательно, скелетные мышцы не могут сокращаться, что приводит к вялому параличу.[10]
Судороги скелетных мышц возникают из-за внезапных и непроизвольных сокращений мышц, которые длятся от секунд до минут и вызывают боль. Хотя они могут быть связаны с заболеваниями, чаще всего мышечные судороги возникают при отсутствии какой-либо явной патологии.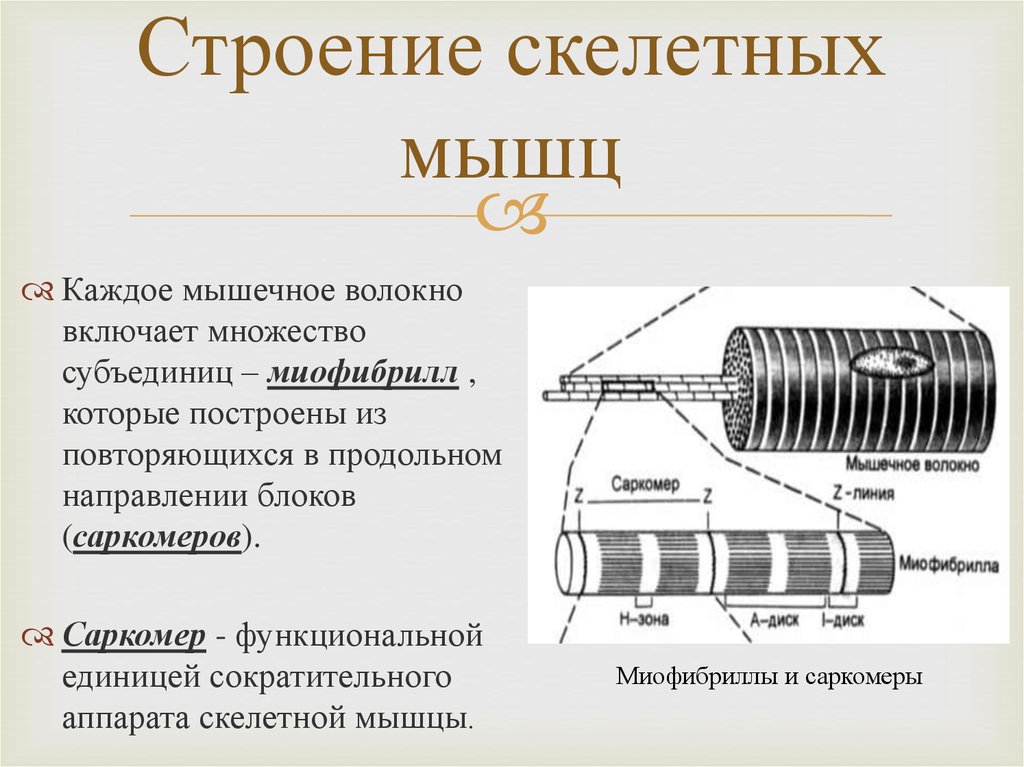 Они чаще всего проявляются у пациентов пожилого возраста, беременных или активно занимающихся физическими упражнениями. Поскольку судороги являются результатом мышечных сокращений, немедленное облегчение боли может быть обеспечено путем растяжения пораженной мышцы.[11]
Они чаще всего проявляются у пациентов пожилого возраста, беременных или активно занимающихся физическими упражнениями. Поскольку судороги являются результатом мышечных сокращений, немедленное облегчение боли может быть обеспечено путем растяжения пораженной мышцы.[11]
Клиническое значение
Оценка мышечной силы и мышечного сокращения является стандартной процедурой, включаемой в физикальное обследование пациента. Шкала ручного мышечного тестирования Совета медицинских исследований является наиболее часто используемой системой оценки мышечной силы, где баллы от 0 до 5 присваиваются в зависимости от способностей пациента. Оценка 0 означает отсутствие активации мышц. Оценка 1 означает, что имеется лишь небольшая сократимость мышцы. 2 балла — это когда активация мышц тестируется в отсутствие гравитации. 3 относится к активации мышц против силы тяжести, но не с сопротивлением. 4 — активация мышц против силы тяжести и некоторого сопротивления, а 5 — активация мышц против гравитации и полного сопротивления. Основная цель проверки мышечной силы при физикальном обследовании состоит в том, чтобы оценить и установить дифференциальный диагноз, когда пациент предъявляет жалобы на слабость, часто на фоне неврологического заболевания. [12]
Основная цель проверки мышечной силы при физикальном обследовании состоит в том, чтобы оценить и установить дифференциальный диагноз, когда пациент предъявляет жалобы на слабость, часто на фоне неврологического заболевания. [12]
Контрольные вопросы
Доступ к бесплатным вопросам с несколькими вариантами ответов по этой теме.
Комментарий к этой статье.
Ссылки
- 1.
Сквайр Дж. Специальный выпуск: Актин-миозиновое взаимодействие в мышцах: история и обзор. Int J Mol Sci. 14 ноября 2019 г. 20 (22) [Бесплатная статья PMC: PMC6887992] [PubMed: 31739584]
- 2.
Оцуки И., Моримото С. Тропонин: регуляторная функция и нарушения. Biochem Biophys Res Commun. 2008 25 апреля; 369(1):62-73. [PubMed: 18154728]
- 3.
Сантулли Г., Льюис Д.Р., Маркс А.Р. Физиология и патофизиология сопряжения возбуждения и сокращения: функциональная роль рианодинового рецептора.
J Muscle Res Cell Motil. 2017 фев;38(1):37-45. [Бесплатная статья PMC: PMC5813681] [PubMed: 28653141]- 4.
Fitts RH. Цикл поперечного моста и усталость скелетных мышц. J Appl Physiol (1985). 2008 г., февраль; 104 (2): 551-8. [PubMed: 18162480]
- 5.
Paolini C, Quarta M, Nori A, Boncompagni S, Canato M, Volpe P, Allen PD, Reggiani C, Protasi F. Реорганизованные запасы и нарушение обработки кальция в скелетных мышцах мышей, лишенных кальсеквестрина-1. Дж. Физиол. 01 сентября 2007 г .: 583 (часть 2): 767–84. [Бесплатная статья PMC: PMC2277031] [PubMed: 17627988]
- 6.
Lieber RL, Ward SR. Конструкция скелетных мышц для удовлетворения функциональных требований. Philos Trans R Soc Lond B Biol Sci. 2011 27 мая; 366 (1570): 1466-76. [Бесплатная статья PMC: PMC3130443] [PubMed: 21502118]
- 7.
Джозефсон Р.К., Эдман К.А. Изменение максимальной скорости укорочения мышечных волокон лягушки в начале тетанического сокращения и во время расслабления.
 J Muscle Res Cell Motil. 2017 фев;38(1):37-45. [Бесплатная статья PMC: PMC5813681] [PubMed: 28653141]
J Muscle Res Cell Motil. 2017 фев;38(1):37-45. [Бесплатная статья PMC: PMC5813681] [PubMed: 28653141]