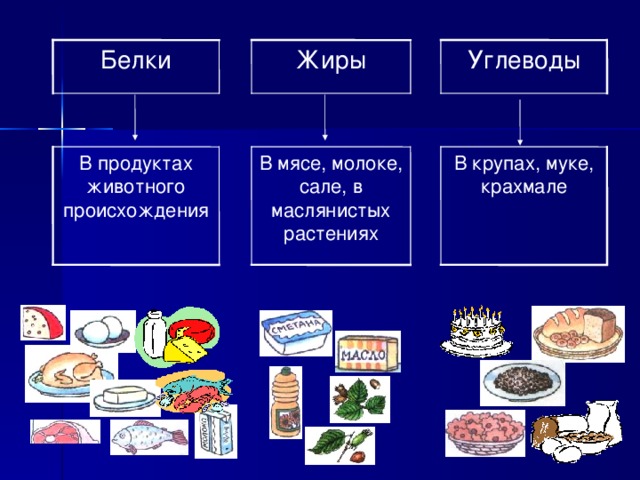
Где содержатся белки и углеводы: Основные принципы здорового питания
Главное, что нужно знать о белках, жирах и углеводах – Sektascience: научно-популярный журнал
У нас есть три большие статьи про белки, жиры и углеводы, но в них много подробностей. Мы написали новую статью с самой важной информацией о БЖУ.
Рассказываем о макронутриентах, их функциях и нормах употребления.
Скачать в PDF
Углеводы — главные поставщики энергии для умственной и физической активности. Различают простые (глюкоза, фруктоза, галактоза, сахароза и др.) и сложные углеводы (крахмалы, пищевые волокна).
| Клетчатка, или пищевые волокна — особая разновидность сложных углеводов, которая почти не усваивается организмом, но дает сытость и помогает поддерживать здоровье желудочно-кишечного тракта. |
Поступая в организм, большая часть углеводов преобразуется в глюкозу, которая используется клетками для жизненных процессов.
Традиционно международные организации здравоохранения рекомендуют потреблять 45–65% углеводов от общей калорийности рациона. Допустимые границы этого диапазона еще шире и зависят от особенностей организма, целей и индивидуальных потребностей.
Как можно чаще отдавайте предпочтение продуктам, содержащим сложные углеводы, — они усваиваются постепенно и не вызывают резких скачков глюкозы, в отличие от простых углеводов. Это важно для поддержания стабильного настроения и предотвращения резких перепадов голода и сытости.
| Основные источники сложных углеводов: крупы, фрукты, овощи, бобовые, картофель, цельнозерновые продукты. |
Простые углеводы содержатся в столовом сахаре, меде и сиропах, выпечке, конфетах и других сладостях, соках и сладких напитках, молочных продуктах.
Белки состоят из соединенных цепочек аминокислот — «строительных» веществ для тела человека.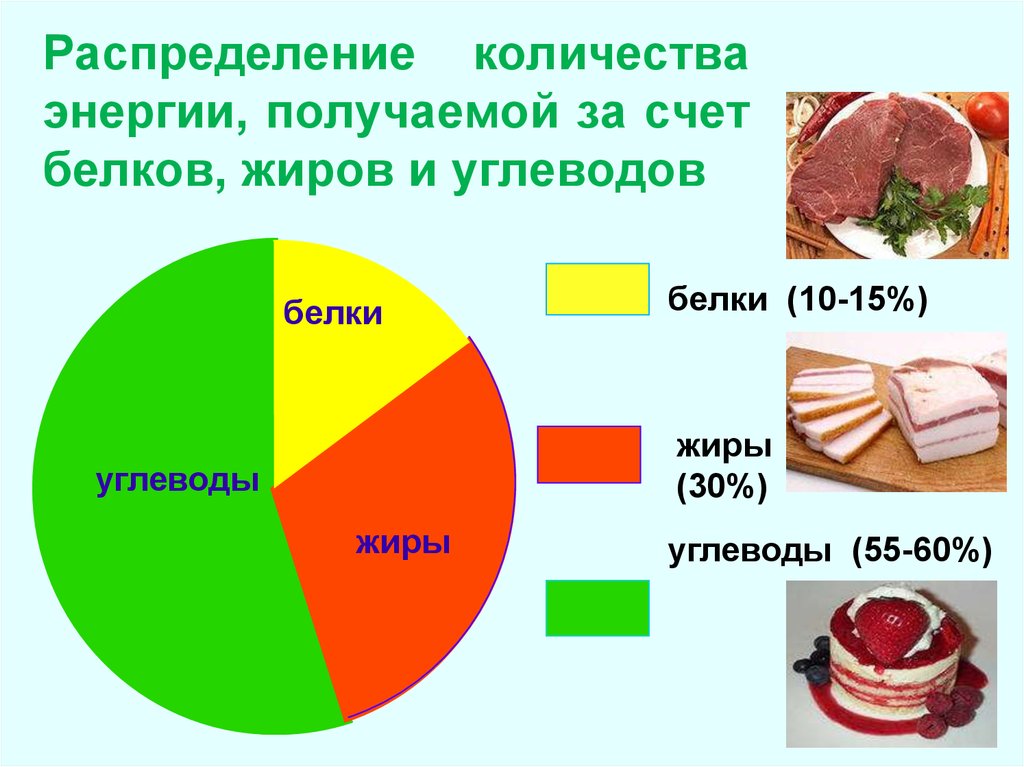 В организме используется двадцать видов аминокислот, девять из которых не могут быть синтезированы телом человека и должны поступать из еды (незаменимые аминокислоты).
В организме используется двадцать видов аминокислот, девять из которых не могут быть синтезированы телом человека и должны поступать из еды (незаменимые аминокислоты).
Все незаменимые аминокислоты содержатся в животном белке. Вегетарианцы тоже могут получить их в полном объеме при сочетании растительных продуктов, богатых белком.
Белки выполняют гормональную, ферментативную и защитную функции в организме. Из них состоят соединительные ткани, мышцы, кожа, волосы и другие органы. Белки не являются прямым источником энергии для организма, но могут быть использованы, когда нет альтернатив или когда усвоение жиров и углеводов нарушено.
Количество белков, необходимых человеку, разнится в среднем от 0,8 до 2,2 г на килограмм веса или 10–35% от общей калорийности рациона. Индивидуальная норма зависит от возраста, пола, активности, периода жизни и других факторов.
Продукты с высоким содержанием белка: мясо и птица, рыба и морепродукты, молоко и молочные продукты, яйца, бобовые и крупы, орехи и семена, соевые продукты. |
Жиры
Жиры — самый энергоемкий макронутриент. Они помогают организму в производстве гормонов, усвоении нутриентов, клеточном росте, защищают от холода и повреждений.
В виде жировой ткани организм может сохранять запасы энергии на долгое время, а в период длительного голодания или низкоуглеводного питания жирные кислоты могут стать основным источником энергии. В этом случае запускается процесс кетоза, с его помощью организм может функционировать с минимальным количеством углеводов.
Именно эта особенность организма помогает людям выживать без еды несколько недель.
Основные виды жиров, или жирных кислот — насыщенные и ненасыщенные.
По данным исследований, чрезмерное потребление насыщенных жиров повышает риск сердечно-сосудистых заболеваний и диабета (но, возможно, это не так).
Основные источники насыщенных жиров — животные продукты (мясо, сливочное масло, сыры и другие молочные продукты, яйца), а также кокосовое и пальмовое масла. |
Употребление ненасыщенных жиров помогает регулировать метаболизм, сохранять эластичность клеточных мембран, улучшает кровообращение, они важны для роста и регенерации клеток.
| Основные источники ненасыщенных жиров : растительные масла, орехи и семена, авокадо, маслины, рыба и птица. |
Отдельно выделяют трансжиры — разновидность ненасыщенных жирных кислот, структура которых делает их устойчивыми к окислению (дольше хранятся и остаются твердыми при комнатной температуре).
В малых количествах трансжиры присутствуют в натуральных мясных и молочных продуктах. В больших — входят в состав некоторых видов маргарина, спредов и кулинарного жира, то есть содержатся в большинстве кондитерских изделий и полуфабрикатов. Часто трансжиры добавляют для увеличения жирности творожных масс, плавленых сырков, мороженого.
Традиционная рекомендация по потреблению жиров: не более 30–35% от общей калорийности рациона, из которых до 10% — насыщенные жиры. Промышленные трансжиры рекомендуется исключать из рациона полностью.
Промышленные трансжиры рекомендуется исключать из рациона полностью.
Все макронутриенты выполняют важные функции в организме человека, однако необходимое соотношение БЖУ в рационе все еще активно обсуждается в научном мире.
Если вы только начали задумываться о том, сколько и чего нужно есть, начните с традиционных рекомендаций организаций здравоохранения о сбалансированном питании. Дальше действуйте по обстоятельствам, ориентируясь на свои цели и самочувствие. Хотите знать больше об этом? У нас есть и подробные статьи:
- Углеводы
- Белки
- Жиры
- Усвоение белков, углеводов и жиров
Важно
- Белки, жиры и углеводы — три основных макронутриента в питании человека, и все они необходимы для функционирования организма и здоровья.
- Все макронутриенты могут служить источниками энергии для организма, но каждый имеет и свои собственные важные функции.
- Большинство продуктов питания содержат больше одного макронутриента, поэтому их невозможно четко поделить на три категории.
 Мы можем только говорить о том, сколько и каких углеводов, белков или жиров приносит каждый продукт в организм.
Мы можем только говорить о том, сколько и каких углеводов, белков или жиров приносит каждый продукт в организм. - Есть только усредненные рекомендованные нормы потребления БЖУ. Индивидуальные потребности в макронутриентах зависят от возраста, пола, активности, периода жизни, целей и других факторов.
- Для обеспечения потребностей организма в макронутриентах выбирайте натуральные, минимально обработанные продукты. Овощи, фрукты, крупы и бобовые богаты клетчаткой и другими сложными углеводами. Те же крупы, бобовые, а также мясо, птица, рыба, морепродукты, яйца, молочные и соевые продукты — источники белка. Рыба, масла, орехи, семена, авокадо и маслины дополнят рацион полезными жирами.
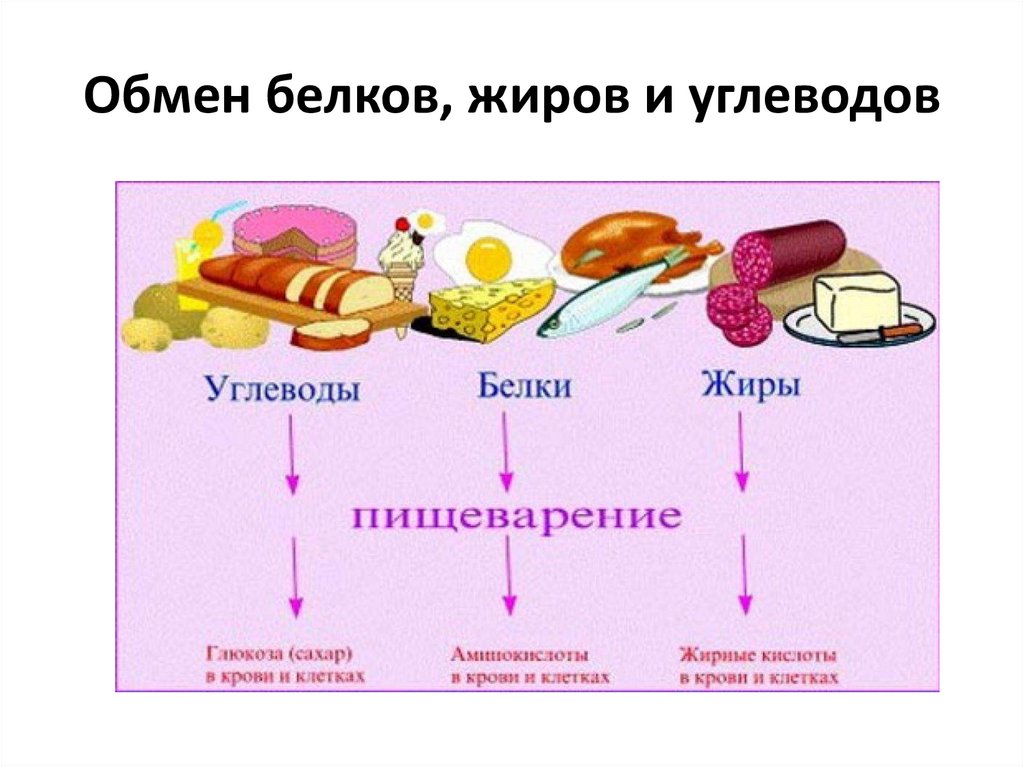 Мы можем только говорить о том, сколько и каких углеводов, белков или жиров приносит каждый продукт в организм.
Мы можем только говорить о том, сколько и каких углеводов, белков или жиров приносит каждый продукт в организм.Автор: Анна Нестерова, тренер по силовым видам спорта,
сотрудник научного отдела Школы идеального тела #sekta
Литература:
1. Dietary guidelines for Americans 2015-2020. (2015.)
2. Types of fat, Harvard Medical School,
3. Trans fat is double trouble for your heart health, Mayo Clinic Staff
4. Mayo Clinic Staff. (2016). Dietary fats: Know which types to choose.
Mayo Clinic Staff. (2016). Dietary fats: Know which types to choose.
5. Saturated, unsaturated, and trans fats. (2018).
6. Annu Rev Nutr. 2006;26:1-22., Fuel metabolism in starvation., Cahill GF Jr(1).
7. The truth about fats: the good, the bad, and the in-between. (2018).
8. World Health Organisation, Topic: Nutrition
Опубликовано в: Питание
В чем польза белков, жиров и углеводов?
Белки, жиры и углеводы – триада компонентов, без которых каждый из нас не мог бы существовать. Их баланс, известных из диетологии, как БЖУ, позволяет составлять оптимальные рационы питания с медицинской и научной точки зрения.
Сами по себе углеводы, жиры и белки является компонентами, входящими в состав всего живого на Земле. Они питают нас, являются материалами для строительства новых клеток и, по сути, являются прямыми составляющими наших с вами тел, участвуя во всех этапах обменных процессов.
Но в чем же польза белков, углеводов и жиров? Давайте разбираться с каждой группой этих веществ по порядку.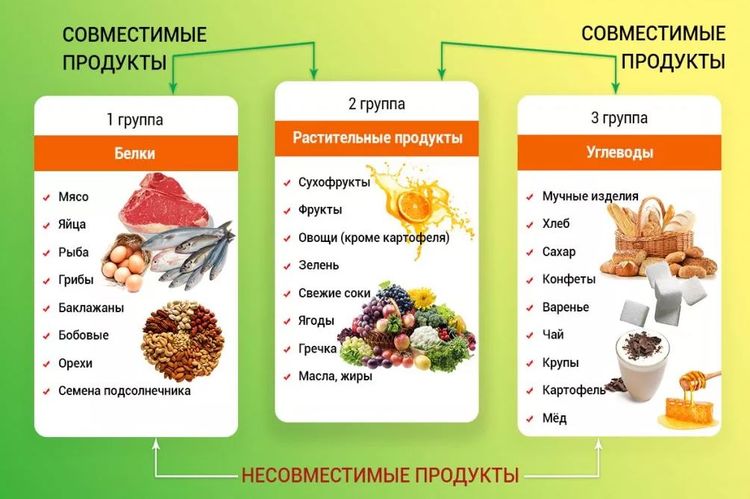
Углеводы и их польза
Углеводы – органические вещества, выполняющие строительную, энергетическую и защитную функцию для организма. Они содержатся во всех фруктах и овощах, а также в популярных в нашей стране крупах и их производных. При этом разделять углеводы принято на сложные и простые. Первые относятся к категории полезных, дающих организму энергию на длительное время и рекомендуемых к регулярному потреблению. Простые, напротив, легко усваиваются и вызывают резкий подъем глюкозы в организме. Это приводит к избытку сахара, получению чрезмерной энергии и образованию излишков жиров, приводящих к избыточному весу.
Именно углеводам уделяется особое внимание при составлении диетического питания. По медицинским показателям снижение их нормы позволяет худеть и поддерживать нужный уровень глюкозы. Однако отказ от углеводов (безуглеводные диеты) могут привести к необратимым болезням печени, гипогликемии и нарушению обменных процессов.
Жиры и их польза
Жиры – вещества, составляющие основу питания всего живого. Они необходимы как для поддержания энергетического баланса, так и для усвоения некоторых групп витаминов и минеральных веществ. При этом многие из нас привыкли ассоциировать жиры с неполезными составляющими питания, пытаясь всячески уменьшить их процент в рационе или попросту отказаться от них.
Они необходимы как для поддержания энергетического баланса, так и для усвоения некоторых групп витаминов и минеральных веществ. При этом многие из нас привыкли ассоциировать жиры с неполезными составляющими питания, пытаясь всячески уменьшить их процент в рационе или попросту отказаться от них.
Жиры необходимы организму. Они являются элементом строительства тканей, транспортерами питательных веществ и энергетическими запасами. Подразделяются они на две большие группы:
- Животные жиры – вещества, получаемые от продуктов животного мира. Они содержат в себе высокий процент холестерина и достаточно медленно усваиваются.
- Растительные жиры – вещества, источником которых служат исключительно растения. Это большинство видов, масел злаковых и семян, авокадо и оливки. Такие жиры усваиваются легче, чем животные, однако не способны в полной мере их заменить.
Белки и их польза
Белки – вещества-строители, выполняющие более 6 важных функций для наших организмов. Они позволяют вырабатывать ферменты, участвуют в процессах транспорта и обмена веществ, являются материалом для строительства клеток, а также отвечают за энергетический баланс.
Они позволяют вырабатывать ферменты, участвуют в процессах транспорта и обмена веществ, являются материалом для строительства клеток, а также отвечают за энергетический баланс.
Основными источниками белков являются мясо и молочная продукция, однако существуют и богатые белком растения. Последние формируют группу растительных белков – все виды бобовых и орехов. Части их используют в постном или вегетарианском меню как альтернативу белкам животным. Однако это не совсем правильно.
Диетологи настоятельно рекомендуют поддерживать должный уровень белков в организме, особенно в период его роста. Вот почему для детского меню существует такое большое количество предписаний, куда в обязательном порядке включаются:
- нежирные сорта рыбы и мяса,
- молоко, йогурт и кефир без ароматических и сладких добавок,
- творог,
- яйца курицы и перепелов.
Структура и функция сложных углеводов — компьютерное моделирование
Сложные углеводы очень распространены в животных, растениях и бактериях. Они входят в состав клеточных мембран, а также субклеточного материала клеток. Они также обнаруживаются в физиологических жидкостях, таких как кровь, слезы, молоко и моча. Недавно было подсчитано, что определены ковалентные структуры от 4000 до 6000 природных углеводов (DOE, 1987). Многие сложные углеводы незамещены по восстанавливающим концам и называются полисахаридами; примеры включают олигосахариды молока, целлюлозу клеточных стенок растений и запасные формы, такие как крахмал и гликоген. Многие другие встречающиеся в природе сложные углеводы ковалентно связаны с другими молекулами, такими как белки или липиды, гликозидными связями остатков сахара на их восстанавливающих концах с образованием гликоконъюгатов.
Они входят в состав клеточных мембран, а также субклеточного материала клеток. Они также обнаруживаются в физиологических жидкостях, таких как кровь, слезы, молоко и моча. Недавно было подсчитано, что определены ковалентные структуры от 4000 до 6000 природных углеводов (DOE, 1987). Многие сложные углеводы незамещены по восстанавливающим концам и называются полисахаридами; примеры включают олигосахариды молока, целлюлозу клеточных стенок растений и запасные формы, такие как крахмал и гликоген. Многие другие встречающиеся в природе сложные углеводы ковалентно связаны с другими молекулами, такими как белки или липиды, гликозидными связями остатков сахара на их восстанавливающих концах с образованием гликоконъюгатов.
БИОЛОГИЧЕСКАЯ ФУНКЦИЯ
Гликопротеины выполняют множество функций в высших организмах. Коллаген является важным структурным элементом внеклеточного пространства, хрящей, костей и базальных мембран. Муцины играют важную роль в качестве смазывающих и защитных агентов в слизистых выделениях. Важные иммунологические молекулы класса гликопротеинов включают иммуноглобулины, антигены гистосовместимости, антигены групп крови типов АВО и Льюиса, комплемент в механизме свертывания крови и интерферон. Многие белки плазмы человека, такие как фетуин, трансферрин и церулоплазмин, являются гликопротеинами, как и некоторые гормоны, такие как хорионический гонадотропин и тиротропин. Большинство животных и растительных лектинов представляют собой гликопротеины, как и лизосомальные ферменты. Распознавание и связывание лизосомальных ферментов со специфическими рецепторами в аппарате Гольджи и на клеточной поверхности включает один или несколько фосфорилированных остатков маннозы на N-связанных олигосахаридных цепях. Сайты узнавания на клеточной поверхности для связывания и поглощения гормонов и для взаимодействия с другими клетками, вирусами и бактериями также являются гликопротеинами.
Важные иммунологические молекулы класса гликопротеинов включают иммуноглобулины, антигены гистосовместимости, антигены групп крови типов АВО и Льюиса, комплемент в механизме свертывания крови и интерферон. Многие белки плазмы человека, такие как фетуин, трансферрин и церулоплазмин, являются гликопротеинами, как и некоторые гормоны, такие как хорионический гонадотропин и тиротропин. Большинство животных и растительных лектинов представляют собой гликопротеины, как и лизосомальные ферменты. Распознавание и связывание лизосомальных ферментов со специфическими рецепторами в аппарате Гольджи и на клеточной поверхности включает один или несколько фосфорилированных остатков маннозы на N-связанных олигосахаридных цепях. Сайты узнавания на клеточной поверхности для связывания и поглощения гормонов и для взаимодействия с другими клетками, вирусами и бактериями также являются гликопротеинами.
Многие функции гликопротеинов на клеточной поверхности также были предложены для нейтральных и кислых гликосфинголипидов. Кроме того, недавно было обнаружено, что некоторые гликосфинголипиды класса ганглиозидов ингибируют митогенный ответ факторов клеточного роста посредством аллостерической модуляции их рецепторов клеточной поверхности (Bremer et al., 1986). Онкогенная трансформация вирусной инфекцией или химическими мутагенами обычно приводит к изменениям структуры гликосфинголипидов на клеточной поверхности, так что количество некоторых типов значительно увеличивается. В некоторых случаях имеются и качественные различия из-за экспрессии генов, молчащих в дифференцированных нормальных клетках. Это особенно важно для опухолевых клеток, где опухолеассоциированные антигены могут служить основой для специфических диагностических анализов на основе моноклональных антител и, в конечном итоге, возможно, для лечения.
Кроме того, недавно было обнаружено, что некоторые гликосфинголипиды класса ганглиозидов ингибируют митогенный ответ факторов клеточного роста посредством аллостерической модуляции их рецепторов клеточной поверхности (Bremer et al., 1986). Онкогенная трансформация вирусной инфекцией или химическими мутагенами обычно приводит к изменениям структуры гликосфинголипидов на клеточной поверхности, так что количество некоторых типов значительно увеличивается. В некоторых случаях имеются и качественные различия из-за экспрессии генов, молчащих в дифференцированных нормальных клетках. Это особенно важно для опухолевых клеток, где опухолеассоциированные антигены могут служить основой для специфических диагностических анализов на основе моноклональных антител и, в конечном итоге, возможно, для лечения.
Связывание между гликозаминогликанами и другими внеклеточными макромолекулами вносит значительный вклад в структурную организацию матрикса соединительной ткани. Все гликозаминогликаны, за исключением тех, в которых отсутствуют сульфатные или карбоксильные группы, электростатически связываются с коллагеном при нейтральном pH из-за их замечательного анионного характера. Дерматансульфат, который, по-видимому, является основным гликозаминогликаном, синтезируемым клетками гладких мышц артерий, сильно связывается с липопротеинами плазмы, а гепарин также взаимодействует с некоторыми белками плазмы, включая факторы свертывания IX и XI и антитромбин III. Интересно, что стехиометрическое связывание гепарина 1:1 с остатками Lys антитромбина III, как полагают, вызывает конформационное изменение антитромбина III, которое увеличивает связывание антитромбина III с тромбином. Это связывание инактивирует тромбин. Гиалуроновая кислота осаждается на поверхности чашек Петри клетками, растущими в культуре тканей, давая им субстрат для прикрепления в процессе роста. Протеогликаны также участвуют в регуляции клеточного роста, возможно, посредством ядерных эффектов на структуру хроматина и активации ДНК-полимеразы, и могут опосредовать межклеточную коммуникацию и потерю рецепторов клеточной поверхности.
Дерматансульфат, который, по-видимому, является основным гликозаминогликаном, синтезируемым клетками гладких мышц артерий, сильно связывается с липопротеинами плазмы, а гепарин также взаимодействует с некоторыми белками плазмы, включая факторы свертывания IX и XI и антитромбин III. Интересно, что стехиометрическое связывание гепарина 1:1 с остатками Lys антитромбина III, как полагают, вызывает конформационное изменение антитромбина III, которое увеличивает связывание антитромбина III с тромбином. Это связывание инактивирует тромбин. Гиалуроновая кислота осаждается на поверхности чашек Петри клетками, растущими в культуре тканей, давая им субстрат для прикрепления в процессе роста. Протеогликаны также участвуют в регуляции клеточного роста, возможно, посредством ядерных эффектов на структуру хроматина и активации ДНК-полимеразы, и могут опосредовать межклеточную коммуникацию и потерю рецепторов клеточной поверхности.
БИОСИНТЕЗ N-СВЯЗАННЫХ ГЛИКОПРОТЕИНОВ И ГЛИКОФИНГОЛИПИДОВ
Роль углеводов в биологических функциях представляет особенно сложную проблему для будущего. Синтез этих гликоконъюгатов происходит во время их внутриклеточного транспорта от места начальной сборки липид-связанного интермедиата (гликопротеины) или церамида (гликосфинголипиды) в эндоплазматическом ретикулуме через аппарат Гольджи на клеточную поверхность, внутриклеточные органеллы или внеклеточные пространство. Для их синтеза требуется семейство активированных доноров сахара, называемых сахарными нуклеотидами, которые синтезируются в цитозольной фракции клеток из сахарофосфатов и нуклеозидтрифосфатов. Интересным исключением является сахарный нуклеотид сиаловой кислоты, называемый цитидинмонофосфат сиаловой кислоты (CMP-NeuAc), который синтезируется в ядре из свободной сиаловой кислоты и CTP. Ферменты, участвующие в биосинтезе гликоконъюгатов, представляют собой гликозилтрансферазы, которые катализируют перенос остатков сахаров от нуклеотидов сахара к невосстанавливающему концу растущей углеводной цепи.
Синтез этих гликоконъюгатов происходит во время их внутриклеточного транспорта от места начальной сборки липид-связанного интермедиата (гликопротеины) или церамида (гликосфинголипиды) в эндоплазматическом ретикулуме через аппарат Гольджи на клеточную поверхность, внутриклеточные органеллы или внеклеточные пространство. Для их синтеза требуется семейство активированных доноров сахара, называемых сахарными нуклеотидами, которые синтезируются в цитозольной фракции клеток из сахарофосфатов и нуклеозидтрифосфатов. Интересным исключением является сахарный нуклеотид сиаловой кислоты, называемый цитидинмонофосфат сиаловой кислоты (CMP-NeuAc), который синтезируется в ядре из свободной сиаловой кислоты и CTP. Ферменты, участвующие в биосинтезе гликоконъюгатов, представляют собой гликозилтрансферазы, которые катализируют перенос остатков сахаров от нуклеотидов сахара к невосстанавливающему концу растущей углеводной цепи.
Различие между биосинтезом гликоконъюгатов и синтезом белка является ключевым; последний происходит на матрице матричной РНК и поэтому определяется генетическим кодом одного структурного гена. 1 В противоположность этому, синтез гликоконъюгатов осуществляется поэтапным добавлением единиц сахара с использованием разных ферментов на каждом этапе. Следовательно, ни одна последовательность ДНК не участвует в определении первичной структуры сложного углевода, поскольку порядок добавления сахаров зависит от специфичности субстрата и кинетических характеристик различных гликозилтрансфераз, каждая из которых кодируется отдельным структурным геном. . Ясно, что невозможно предсказать первичную структуру сложных углеводов по последовательностям ДНК. Следовательно, трехмерные структуры гликопротеинов, гликосфинголипидов и других сложных молекул, содержащих углеводы, невозможно полностью предсказать без экспериментального структурного анализа углеводов.
1 В противоположность этому, синтез гликоконъюгатов осуществляется поэтапным добавлением единиц сахара с использованием разных ферментов на каждом этапе. Следовательно, ни одна последовательность ДНК не участвует в определении первичной структуры сложного углевода, поскольку порядок добавления сахаров зависит от специфичности субстрата и кинетических характеристик различных гликозилтрансфераз, каждая из которых кодируется отдельным структурным геном. . Ясно, что невозможно предсказать первичную структуру сложных углеводов по последовательностям ДНК. Следовательно, трехмерные структуры гликопротеинов, гликосфинголипидов и других сложных молекул, содержащих углеводы, невозможно полностью предсказать без экспериментального структурного анализа углеводов.
Snider (1984) сообщил, что гликопротеины N-связанного типа синтезируются как котрансляционное событие в шероховатой эндоплазматической сети. В то время как полипептидная цепь транслируется на матричной РНК и одновременно проходит через мембрану эндоплазматического ретикулума в цистернальное пространство (просвет), одиночный олигосахарид координированно синтезируется на фосфорилированном полиизопреноидном спирте (долихоле у высших животных и более мелких подобных веществах у насекомых).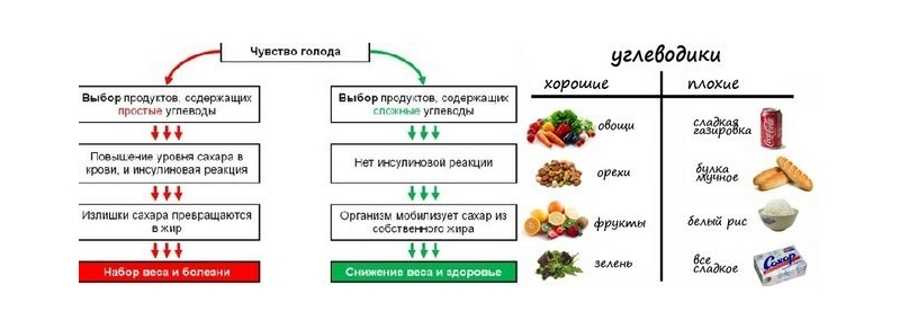 , дрожжи и растения). Весь олигосахарид-предшественник затем переносится на соответствующие остатки аспарагина в формирующейся полипептидной цепи (вероятно, перед укладкой в третичную структуру) в соответствии с правилами специфичности, которые до конца не поняты. Для переноса требуется последовательность Asn-X-Ser или Asn-X-Thr, но также задействованы дополнительные факторы. Доступность остатка Asn может быть одним из таких факторов, и оценка этой возможности может быть сделана с помощью методов прогнозирования, описанных в этом отчете.
, дрожжи и растения). Весь олигосахарид-предшественник затем переносится на соответствующие остатки аспарагина в формирующейся полипептидной цепи (вероятно, перед укладкой в третичную структуру) в соответствии с правилами специфичности, которые до конца не поняты. Для переноса требуется последовательность Asn-X-Ser или Asn-X-Thr, но также задействованы дополнительные факторы. Доступность остатка Asn может быть одним из таких факторов, и оценка этой возможности может быть сделана с помощью методов прогнозирования, описанных в этом отчете.
Вторая стадия синтеза N-связанного гликопротеина включает обширную посттрансляционную модификацию связанного с белком олигосахарида-предшественника путем удаления и добавления сахаров. Во многих случаях белковая часть также модифицируется частичным протеолитическим расщеплением и/или добавлением модифицирующих функцию групп к определенным аминокислотным остаткам. Посттрансляционная модификация инициируется в шероховатой эндоплазматической сети удалением трех остатков глюкозы двумя специфическими мембраносвязанными глюкозидазами. Эти остатки глюкозы, по-видимому, имеют единственную функцию обеспечения переноса олигосахаридной цепи от долихолпирофосфата к зарождающимся полипептидным цепям. Будет интересно определить по трехмерным структурам и предсказанным конформациям, как эти группы взаимодействуют с ферментом трансферазой, задействованным на этом этапе. Цепи зрелых олигосахаридов с высоким содержанием маннозы синтезируют путем последующего удаления до четырех остатков маннозила из трех ветвей структуры предшественника. В этом процессе участвуют по крайней мере три различных альфа-маннозидазы в аппарате Гольджи. Эти ферменты и две глюкозидазы являются гидролазами, как и лизосомальные гликозидазы, но их активность максимальна при нейтральном рН, в отличие от лизосомальных ферментов, которые проявляют наибольшую каталитическую активность при кислом рН.
Эти остатки глюкозы, по-видимому, имеют единственную функцию обеспечения переноса олигосахаридной цепи от долихолпирофосфата к зарождающимся полипептидным цепям. Будет интересно определить по трехмерным структурам и предсказанным конформациям, как эти группы взаимодействуют с ферментом трансферазой, задействованным на этом этапе. Цепи зрелых олигосахаридов с высоким содержанием маннозы синтезируют путем последующего удаления до четырех остатков маннозила из трех ветвей структуры предшественника. В этом процессе участвуют по крайней мере три различных альфа-маннозидазы в аппарате Гольджи. Эти ферменты и две глюкозидазы являются гидролазами, как и лизосомальные гликозидазы, но их активность максимальна при нейтральном рН, в отличие от лизосомальных ферментов, которые проявляют наибольшую каталитическую активность при кислом рН.
В эукариотических клетках олигосахарид с высоким содержанием маннозы с пятью единицами маннозы (см. ) является прямым предшественником сложных и гибридных структур. Начальным этапом в аппарате Гольджи является добавление остатка N-ацетилглюкозамина к последнему оставшемуся человеку на ветви I (*), после чего оставшиеся два остатка человека на ветвях II и III могут быть удалены альфа-маннозидазами, которые почти определенно отличается от тех, кто участвовал в предыдущих шагах. На этом этапе могут быть сделаны дополнительные разветвления для получения три- и тетраантеннальных структур, а заключительные стадии процессинга осуществляются путем добавления остатков галактозы, N-ацетилглюкозамина, сиаловой кислоты и фукозы для получения зрелых, сложных, N-связанных цепи. Интересная N-ацетилглюкозаминилтрансфераза может добавлять бета-1,4-связанный остаток GlcNAc к разветвленному бета-связанному остатку маннозы внутренней области ядра (0), чтобы получить «разделенную пополам структуру». Этот этап был предметом интенсивного изучения Карвером и его коллегами, которые интересовались структурной специфичностью фермента с различными конформациями предшественников олигосахаридов (Карвер и Бриссон, 19).
Начальным этапом в аппарате Гольджи является добавление остатка N-ацетилглюкозамина к последнему оставшемуся человеку на ветви I (*), после чего оставшиеся два остатка человека на ветвях II и III могут быть удалены альфа-маннозидазами, которые почти определенно отличается от тех, кто участвовал в предыдущих шагах. На этом этапе могут быть сделаны дополнительные разветвления для получения три- и тетраантеннальных структур, а заключительные стадии процессинга осуществляются путем добавления остатков галактозы, N-ацетилглюкозамина, сиаловой кислоты и фукозы для получения зрелых, сложных, N-связанных цепи. Интересная N-ацетилглюкозаминилтрансфераза может добавлять бета-1,4-связанный остаток GlcNAc к разветвленному бета-связанному остатку маннозы внутренней области ядра (0), чтобы получить «разделенную пополам структуру». Этот этап был предметом интенсивного изучения Карвером и его коллегами, которые интересовались структурной специфичностью фермента с различными конформациями предшественников олигосахаридов (Карвер и Бриссон, 19). 84).
84).
РИСУНОК 8-1
Промежуточный продукт, частично процессированный аспарагином — связанная углеводная цепь гликопротеина.
Вполне вероятно, что в исследованиях путей процессинга и степени процессинга цепей олигосахаридов будут использоваться методы прогнозирования. Если контроль возникает из-за специфичности фермента в отношении конкретной трехмерной структуры субстрата, может оказаться возможным определить эти предпочтения и, исходя из предсказаний распределения трехмерных структур олигосахарида, присоединенного к гликопротеиновому субстрату, предсказать, насколько далеко углеводная цепь будет обработана.
Лизосомальные ферменты содержат одну или несколько фосфатных групп на маннозных остатках олигосахаридных цепей с высоким содержанием маннозы. Маннозо-6-фосфатные группы являются специфическими маркерами распознавания, которые участвуют в транспорте лизосомальных ферментов из аппарата Гольджи или из клеток в лизосомы. В плазматической мембране были обнаружены два мембраносвязанных маннозо-6-фосфатных рецептора; по крайней мере, один из них также находится в мембранах Гольджи.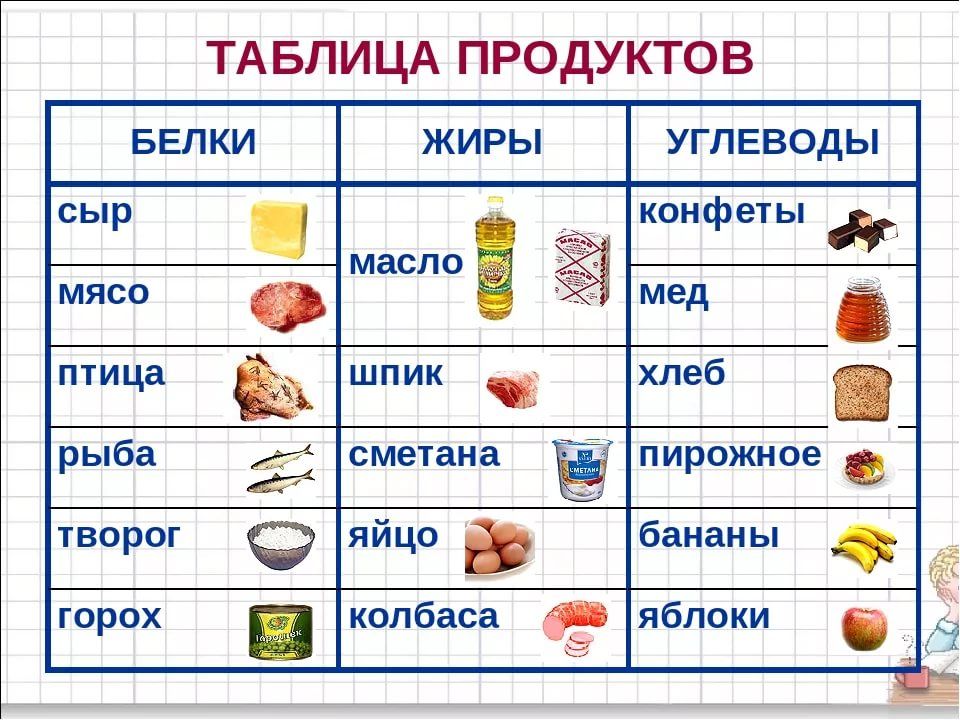 Хотя специфичность их связывания исследована довольно подробно, другие аспекты не определены: природа взаимодействия фосфорилированных остатков маннозы с рецепторами и трехмерные структуры лизосомальных фермент-рецепторных комплексов.
Хотя специфичность их связывания исследована довольно подробно, другие аспекты не определены: природа взаимодействия фосфорилированных остатков маннозы с рецепторами и трехмерные структуры лизосомальных фермент-рецепторных комплексов.
Другой интересный аспект синтеза лизосомальных ферментов включает определение структурных доменов на свернутых белках, распознаваемых ферментом, который инициирует фосфорилирование остатков маннозы, который представляет собой N-ацетилглюкозамин-фосфотрансферазу (GlcNAc-P трансферазу) в аппарате Гольджи. Это механизм, с помощью которого для фосфорилирования отбираются только белки лизосомальных ферментов. Это особенно важно, поскольку одна из форм генетического лизосомного нарушения накопления, называемая муколипидозом II, возникает в результате дефекта связывающего домена GlcNAc-P трансферазы для лизосомальных ферментных белков. Возможно, эту проблему можно решить только с помощью компьютерного моделирования, позволяющего предсказать трехмерные структуры обоих белков.
Гликосфинголипиды синтезируются аналогичным образом, за исключением того, что церамид выполняет функцию долихола для гликопротеинов, а перенос происходит непосредственно от сахарного нуклеотида к акцепторному гликолипиду. Церамид является акцептором либо глюкозы (из UDP-Glc), либо галактозы (из UDP-Gal), образуя глюкозилцерамид или галактозилцерамид. Эти простые гликосфинголипиды преобладают в плазме и головном мозге человека соответственно, а также служат предшественниками более сложных гликосфинголипидов. В большинстве органов, включая головной мозг, основные пути включают превращение глюкозилцерамида в лактозилцерамид, Gal-бета-1,4-Glc-Cer. Лактозилцерамид является субстратом ряда гликозилтрансфераз, продукты которых являются первыми интермедиатами в синтезе родственных гликосфинголипидов, которые можно классифицировать по их общим структурным характеристикам. Уже охарактеризовано более 100 различных гликосфинголипидов, и все еще открываются новые соединения. Хотя некоторые из гликосфинголипидов могут содержать от 15 до 35 или более остатков сахара, большинство широко встречающихся типов имеют от 4 до 10 остатков в олигосахаридной цепи.
АНАЛИЗ ПЕРВИЧНОЙ И ТРЕТИЧНОЙ СТРУКТУРЫ
Полное понимание взаимодействий между углеводами и белками (ферментами, лектинами, антителами и рецепторами клеточной поверхности) будет зависеть от определения точных трехмерных структур обоих типов молекул. Как было отмечено, первичные структуры олигосахаридных цепей сложных углеводов не могут быть выведены из последовательностей ДНК и поэтому должны быть определены химическим и спектроскопическим анализом. Современные хроматографические методы разделения, наряду с масс-спектрометрией и ядерным магнитным резонансом (ЯМР), позволяют проводить полный анализ первичной структуры на образце в один микромоль. Еще предстоит определить состав; размещение остатков сахара; размер кольца; положения гликозидных связей и их аномерность; и расположение и химическая природа неуглеводных заместителей, таких как липиды, сульфатные и фосфатные группы.
Трехмерные структуры углеводов представляют пространственное расположение отдельных остатков сахара. Наиболее часто встречающиеся сложные углеводы млекопитающих состоят из остатков сахаров, которые существуют в форме пиранозного кольца, наиболее стабильной и жесткой конформацией которого являются формы стула. Когда два остатка сахара ковалентно соединены гликозидной связью, они могут свободно вращаться вокруг гликозидного атома кислорода между двумя кольцами, и поэтому полученный дисахарид может принимать ряд различных конформаций, соответствующих вращениям вокруг этих двух связей. Двугранные углы при гликозидной связи (см. ) принято обозначать греческими символами phi (ϕ) и psi (ψ), где исходной конформацией (ϕ = 0°, ψ = 0°) является тот конформер, в котором C-l Связь -H-1 затмевает O-C’-X’, а связь C-1-O затмевает C’-X’-H-X’.
Наиболее часто встречающиеся сложные углеводы млекопитающих состоят из остатков сахаров, которые существуют в форме пиранозного кольца, наиболее стабильной и жесткой конформацией которого являются формы стула. Когда два остатка сахара ковалентно соединены гликозидной связью, они могут свободно вращаться вокруг гликозидного атома кислорода между двумя кольцами, и поэтому полученный дисахарид может принимать ряд различных конформаций, соответствующих вращениям вокруг этих двух связей. Двугранные углы при гликозидной связи (см. ) принято обозначать греческими символами phi (ϕ) и psi (ψ), где исходной конформацией (ϕ = 0°, ψ = 0°) является тот конформер, в котором C-l Связь -H-1 затмевает O-C’-X’, а связь C-1-O затмевает C’-X’-H-X’.
РИСУНОК 8-2
Двугранные углы, определяющие пространственное отношение двух остатков сахара в дисахариде.
Относительная ориентация соседних сахарных остатков в олигосахаридной цепи описывается указанием углов вращения (ϕ, ψ) у каждого гликозидного атома кислорода.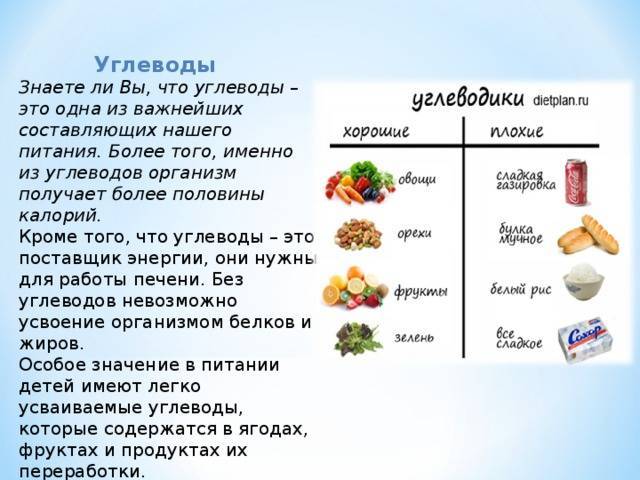 Когда эти углы одинаковы для каждой связи, цепь имеет спиральную конформацию с n остатками на виток и h единицами перемещения вдоль оси спирали. Если n и h доступны из рентгеновских данных, то можно вычислить ϕ и ψ и наоборот. Если ϕ и ψ различны среди гликозидных связей в олигосахаридной цепи, трехмерная структура становится непериодической и, при крайних вариациях, принимает конформацию случайного клубка. Информация о возмущениях может быть получена с помощью измерений светорассеяния, вязкости, седиментации и диффузии.
Когда эти углы одинаковы для каждой связи, цепь имеет спиральную конформацию с n остатками на виток и h единицами перемещения вдоль оси спирали. Если n и h доступны из рентгеновских данных, то можно вычислить ϕ и ψ и наоборот. Если ϕ и ψ различны среди гликозидных связей в олигосахаридной цепи, трехмерная структура становится непериодической и, при крайних вариациях, принимает конформацию случайного клубка. Информация о возмущениях может быть получена с помощью измерений светорассеяния, вязкости, седиментации и диффузии.
РЕНТГЕНОВСКИЙ АНАЛИЗ КРИСТАЛЛИЧЕСКИХ СТРУКТУР УГЛЕВОДОВ
Из трех основных классов сложных биологических молекул у нас есть наименьшая структурная информация об углеводах с атомным разрешением. Это связано с тем, что они не были кристаллизованы, и, следовательно, нет соответствующей базы данных о кристаллической структуре, кроме базы данных от простых мономеров до тримеров, на основе которой можно было бы моделировать классические или полуэмпирические квантово-механические расчеты. Типичными примерами являются олигосахариды, специфичные для группы крови, кордовые факторы и липиды A и X. Исключением являются циклодекстрины, которые хорошо кристаллизуются, но конформационно относятся к отдельному классу. Структуры, полученные из структуры волокон полисахаридов, зависят от модели и не являются источником окончательных структурных данных. Стахиоза, олигосахарид, состоящий из четырех сахарных остатков, является крупнейшим нециклическим олигосахаридом, для которого проводится анализ кристаллической структуры, но даже в этом случае связанная с ним структура воды не определена.
Типичными примерами являются олигосахариды, специфичные для группы крови, кордовые факторы и липиды A и X. Исключением являются циклодекстрины, которые хорошо кристаллизуются, но конформационно относятся к отдельному классу. Структуры, полученные из структуры волокон полисахаридов, зависят от модели и не являются источником окончательных структурных данных. Стахиоза, олигосахарид, состоящий из четырех сахарных остатков, является крупнейшим нециклическим олигосахаридом, для которого проводится анализ кристаллической структуры, но даже в этом случае связанная с ним структура воды не определена.
Проблема кристалличности присуща только частично. Углеводы сольватируются иначе, чем белки, олигонуклеотиды или нуклеиновые кислоты. Однако для получения значительных количеств конфигурационно-однородного материала , необходимого для проведения экспериментов по кристаллизации, было предпринято меньше усилий, чем для белков и нуклеиновых кислот. Другим аспектом кристаллографии гликоконъюгатов является то, что электронная плотность для олигосахаридной части гликопротеинов редко интерпретируется, даже несмотря на то, что было изучено несколько кристаллических гликопротеинов.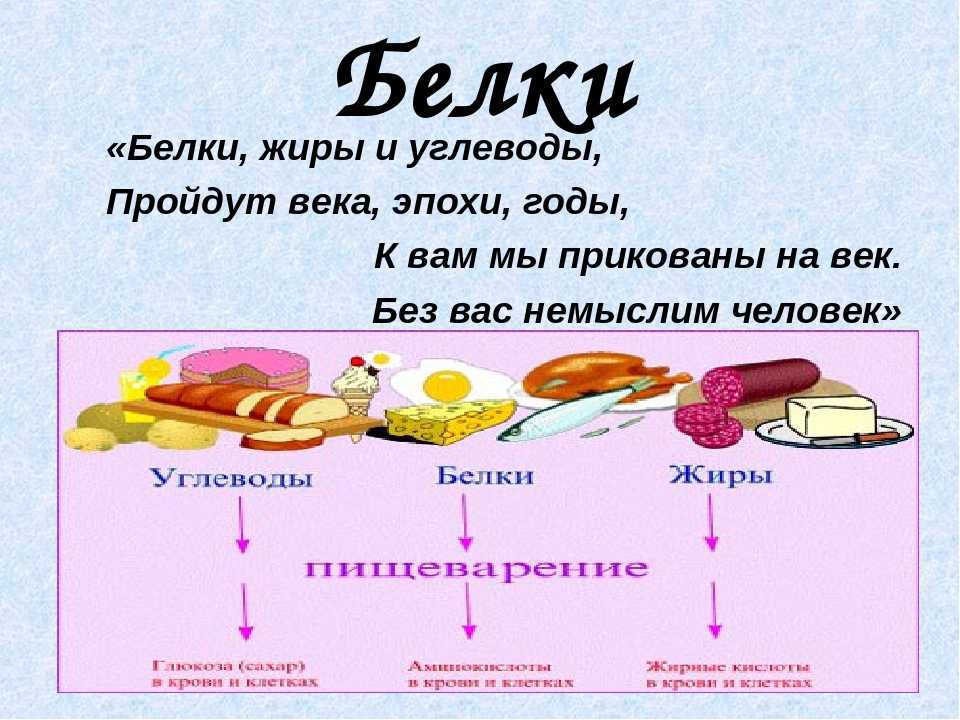 Это связано с тем, что стандартные программы уточнения не могут работать с олигосахаридами или в месте гликозилирования присутствует микрогетерогенность, поэтому он не учитывается в модели. Таким образом, потенциально ценный источник информации не используется из-за отсутствия надлежащей разработки программ или стратегических подходов к решению проблемы микрогетерогенности.
Это связано с тем, что стандартные программы уточнения не могут работать с олигосахаридами или в месте гликозилирования присутствует микрогетерогенность, поэтому он не учитывается в модели. Таким образом, потенциально ценный источник информации не используется из-за отсутствия надлежащей разработки программ или стратегических подходов к решению проблемы микрогетерогенности.
Стерические соображения о минимальных расстояниях сближения между атомами, полученные из наблюдаемых несвязанных расстояний в различных кристаллических структурах, могут быть использованы для предсказания допустимых конформаций. Этот подход «твердой сферы», первоначально разработанный В.С.Р. Рао в середине 1970-х годов, представляет собой рудиментарный метод теоретических расчетов, который игнорирует электростатические эффекты (водородные связи), но дает качественное предсказание структуры. Впоследствии этот подход был расширен за счет адаптации расчетов энергии, первоначально использовавшихся для пептидов, где потенциальная энергия делится на функции, описывающие дискретные вклады, такие как энергии Ван-дер-Ваальса, электростатические взаимодействия, энергия кручения, энергия водородных связей, деформации связей и углов (Бок , 1983). Данные представлены в виде компьютерных карт энергетических контуров.
Данные представлены в виде компьютерных карт энергетических контуров.
В большей части недавней литературы расчеты конформационной энергии проводились с использованием формы параметров Рао с добавленным торсионным потенциалом относительно одной из гликозидных связей (экзоаномерный эффект). Этот подход, получивший название HSEA (экзоаномерный метод твердых сфер) (Bock, 1983), с успехом использовался Лемье и Боком (1983), Карвером и Бриссоном (1984) и другими, хотя он содержит ряд непроверенных предположений. Добавление потенциала водородной связи (метод HEAH) дает результаты минимизации энергии, которые отличаются от результатов, рассчитанных методом HSEA, из которых можно получить геометрию, отличающуюся от геометрии, полученной методом HSEA.
ЯМР РАСТВОРЫ СТРУКТУРЫ УГЛЕВОДОВ
Методы протонного ЯМР предоставляют подробные экспериментальные данные, на основании которых можно определить трехмерные структуры и сравнить их с конформациями, полученными расчетами потенциальной энергии.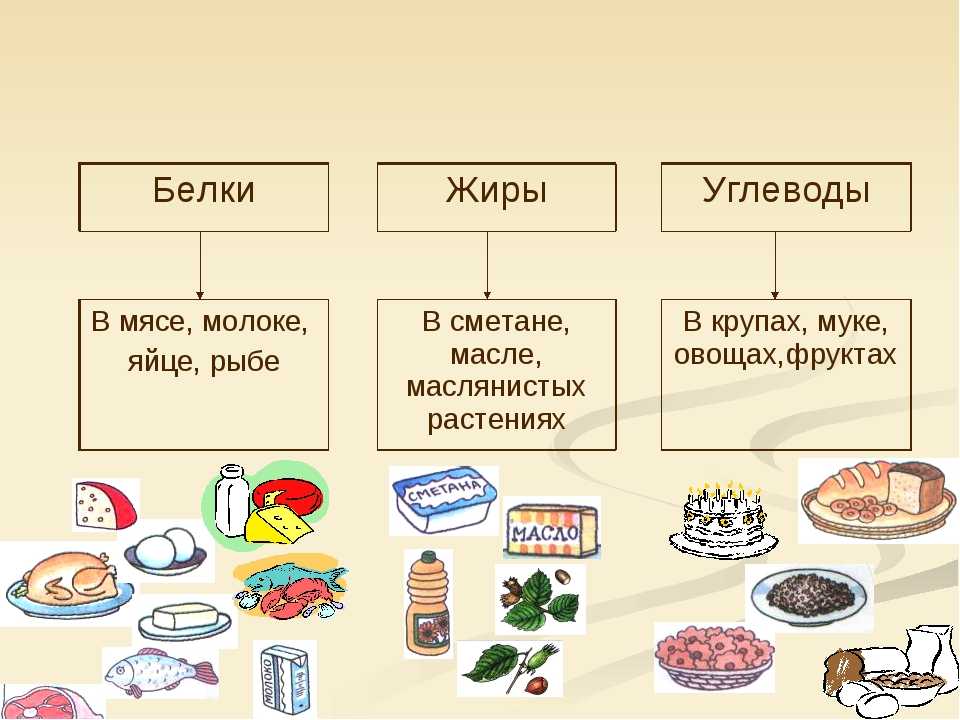 Карвер и Камминг (1987) создали контурные карты рассчитанных NOE различных олигосахаридов с высоким содержанием маннозы в зависимости от торсионных углов φ и ψ. Затем они связали их с экспериментальными результатами, а также с конформациями минимальной энергии, оцененными с помощью различных расчетов потенциальной энергии (Карвер и Камминг, в печати). Бриссон и Карвер (1983) оценил полезность этого подхода с использованием двух гликопептидов биантеннарного сложного типа (см. ). Поскольку конформации, полученные с помощью NOE, находились в диапазоне, центрированном на конформациях с минимальной энергией, полученных из расчетов потенциальной энергии, был сделан вывод, «что усреднение движения ограничено узким диапазоном около одной стабильной конформации» (Brisson and Carver, 1983). Однако теперь оказывается, что бессмысленно искать единственную конформацию, полученную из NOE, которая удовлетворяет одному минимуму потенциальной энергии, потому что фактически молекулы могут занимать такие минимумы в течение очень небольшой части времени в растворе.
Карвер и Камминг (1987) создали контурные карты рассчитанных NOE различных олигосахаридов с высоким содержанием маннозы в зависимости от торсионных углов φ и ψ. Затем они связали их с экспериментальными результатами, а также с конформациями минимальной энергии, оцененными с помощью различных расчетов потенциальной энергии (Карвер и Камминг, в печати). Бриссон и Карвер (1983) оценил полезность этого подхода с использованием двух гликопептидов биантеннарного сложного типа (см. ). Поскольку конформации, полученные с помощью NOE, находились в диапазоне, центрированном на конформациях с минимальной энергией, полученных из расчетов потенциальной энергии, был сделан вывод, «что усреднение движения ограничено узким диапазоном около одной стабильной конформации» (Brisson and Carver, 1983). Однако теперь оказывается, что бессмысленно искать единственную конформацию, полученную из NOE, которая удовлетворяет одному минимуму потенциальной энергии, потому что фактически молекулы могут занимать такие минимумы в течение очень небольшой части времени в растворе.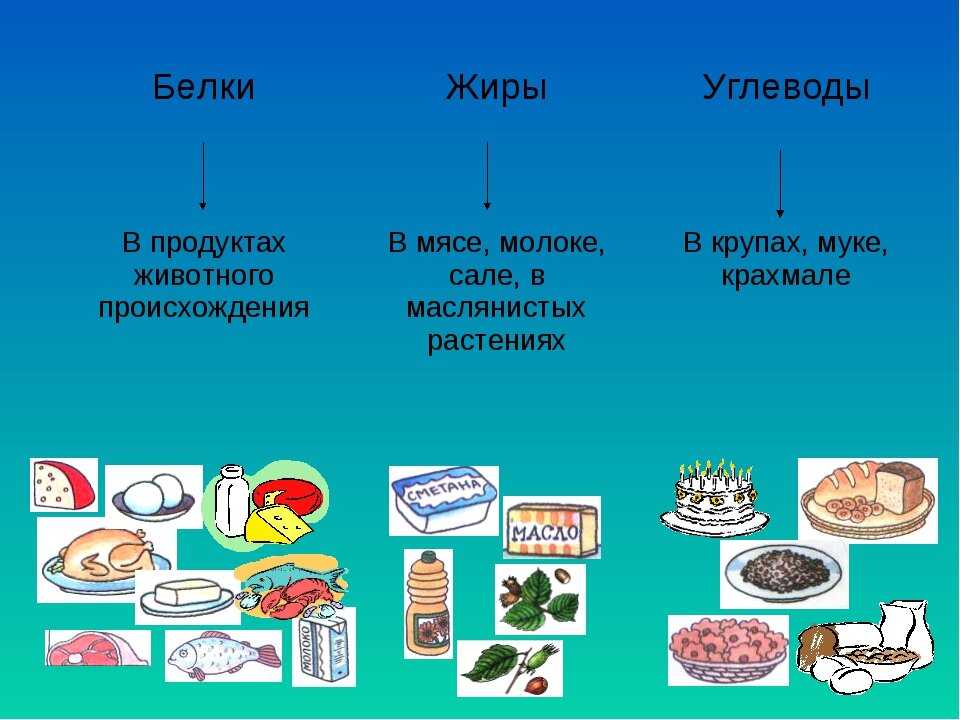 «Конформационная гибкость должна быть включена в теоретическую трактовку» (Карвер и Камминг, 19).87), и расчет энергетических поверхностей становится чрезвычайно важным. Последние исследования Камминга и Карвера показывают, что трехмерные структуры, определяемые NOE, могут значительно отличаться от любой конформации с минимальной энергией. Из этого они пришли к выводу, что конформации, полученные из NOE, в таких случаях могут соответствовать «виртуальным» конформациям, как определено Jardetzky (1980), как вычисленные структуры, которые на самом деле принимают лишь немногие молекулы в растворе, если вообще какие-либо.
«Конформационная гибкость должна быть включена в теоретическую трактовку» (Карвер и Камминг, 19).87), и расчет энергетических поверхностей становится чрезвычайно важным. Последние исследования Камминга и Карвера показывают, что трехмерные структуры, определяемые NOE, могут значительно отличаться от любой конформации с минимальной энергией. Из этого они пришли к выводу, что конформации, полученные из NOE, в таких случаях могут соответствовать «виртуальным» конформациям, как определено Jardetzky (1980), как вычисленные структуры, которые на самом деле принимают лишь немногие молекулы в растворе, если вообще какие-либо.
РИСУНОК 8-3
Структуры двух частично процессированных аспарагиновых связанных углеводных цепей. Разделение пополам β1,4GlcNAc B вызывает конформационное отличие от A.
Scarsdale et al. (в печати) использовали программу, основанную на молекулярной механике, в попытке смоделировать конформационное усреднение данных ЯМР. Конформации рассчитывали с использованием комбинации молекулярных потенциалов и данных ЯМР для олигосахаридной части эритроцитарного гликолипида, состоящего из трех нейтральных сахаров и аминосахара. Конформер с наименьшей энергией очень напоминал структуру, предложенную ранее. Однако соответствие данным можно улучшить, если рассмотреть два уравновешивающих конформера. Таким образом, можно определить конформации растворов сложных углеводов даже в нежестких случаях, используя комбинацию расчетов и ограничений, налагаемых экспериментальными данными ЯМР.
Конформер с наименьшей энергией очень напоминал структуру, предложенную ранее. Однако соответствие данным можно улучшить, если рассмотреть два уравновешивающих конформера. Таким образом, можно определить конформации растворов сложных углеводов даже в нежестких случаях, используя комбинацию расчетов и ограничений, налагаемых экспериментальными данными ЯМР.
Несмотря на поднятые вопросы об интерпретации результатов ЯМР и ценности потенциальной минимизации энергии, была собрана некоторая важная информация о взаимодействиях углеводных антигенов с антителами (Lemieux et al., 1985), олигосахаридов с лектинами, такими как конканавалин А ( Sekharudu et al., 1986), и олигосахариды с ферментами гликозилтрансферазами (Carver and Cumming, 1987). Дальнейшие уточнения будут зависеть от разработки согласованного набора функций потенциальной энергии, которые можно использовать с экспериментально определенными трехмерными структурами, полученными из NOE, для оценки того, распределяется ли данная молекула между несколькими низкоэнергетическими конформациями или занимает конкретную форму.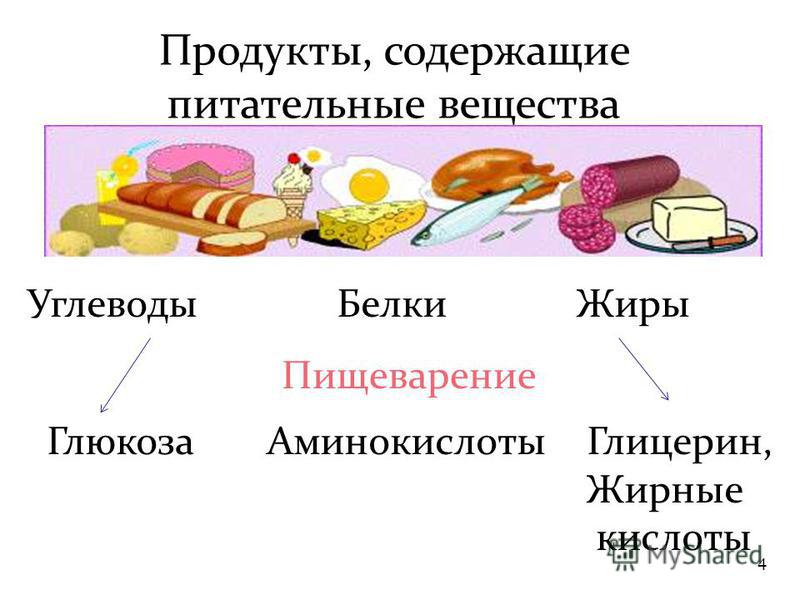 их подмножество. Твароска и Перес (1986) недавно сравнили несколько расчетов конформационной энергии и предложили общую стратегию для олигосахаридов.
их подмножество. Твароска и Перес (1986) недавно сравнили несколько расчетов конформационной энергии и предложили общую стратегию для олигосахаридов.
Компьютерное время и доступ к соответствующим процессорам массива параллельной обработки являются важными соображениями при определении уровня поддержки исследований в этой области в настоящее время. Доступность машин для очень быстрого расчета межатомных расстояний и вкладов Ван-дер-Ваальса — это вопрос, который должны решать финансирующие агентства. Интересно, что несколько суперкомпьютеров, работающих в настоящее время в кампусах, не используются на полную мощность; возможно, усилия соответствующих консультативных групп в этих центрах должны быть направлены на разработку необходимого программного обеспечения для этих компьютеров и установление политики, которая будет направлять часть их времени на компьютерное моделирование трехмерных структур.
НАДМОЛЕКУЛЯРНАЯ СТРУКТУРА
Структуры, состоящие из более чем одной макромолекулы, взаимодействуют как единое целое в биологических явлениях, таких как катализ многими ферментами, связывание на клеточной поверхности, передача сигнала через клеточные мембраны и другие биологические явления. Любой фермент, состоящий более чем из одной субъединицы, следует рассматривать как надмолекулярную структуру. Когда задействовано большое количество субъединиц и, возможно, они выполняют более одной функции, может потребоваться особое внимание к их относительной пространственной ориентации. Примерами являются репликация ДНК с помощью ДНК-полимераз, когда для инициации репликации требуются комплексы, содержащие 10 или 12 белков (называемые примосомами). Рибосомы еще более сложны: для трансляции матричной РНК требуется не менее 75 белков. Поверхности, состоящие более чем из одной макромолекулы, часто ведут себя как функциональная единица. Например, поглощение холестерина многими клетками требует взаимодействия специфического рецептора клеточной поверхности с полипептидной поверхностью сложной надмолекулярной структуры, называемой липопротеином низкой плотности (ЛПНП), который состоит из белка, холестерина, фосфолипидов и триацилглицеролов. Изменение белка ЛПНП путем ацетилирования остатка Lys блокирует связывание ЛПНП с его рецептором и поглощение холестерина клеткой.
Любой фермент, состоящий более чем из одной субъединицы, следует рассматривать как надмолекулярную структуру. Когда задействовано большое количество субъединиц и, возможно, они выполняют более одной функции, может потребоваться особое внимание к их относительной пространственной ориентации. Примерами являются репликация ДНК с помощью ДНК-полимераз, когда для инициации репликации требуются комплексы, содержащие 10 или 12 белков (называемые примосомами). Рибосомы еще более сложны: для трансляции матричной РНК требуется не менее 75 белков. Поверхности, состоящие более чем из одной макромолекулы, часто ведут себя как функциональная единица. Например, поглощение холестерина многими клетками требует взаимодействия специфического рецептора клеточной поверхности с полипептидной поверхностью сложной надмолекулярной структуры, называемой липопротеином низкой плотности (ЛПНП), который состоит из белка, холестерина, фосфолипидов и триацилглицеролов. Изменение белка ЛПНП путем ацетилирования остатка Lys блокирует связывание ЛПНП с его рецептором и поглощение холестерина клеткой.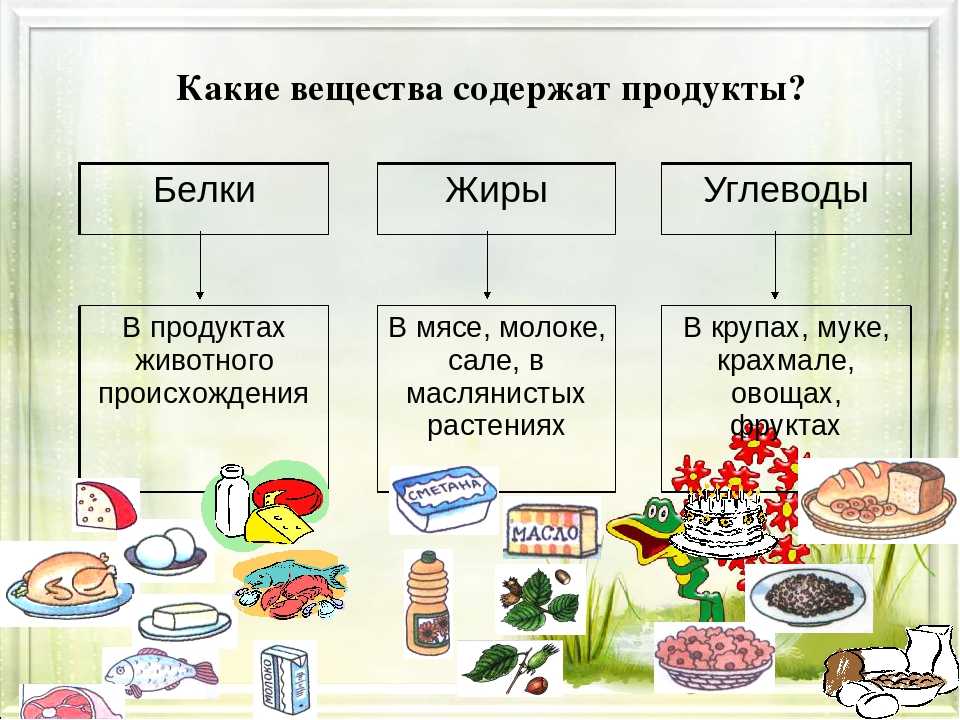 Некоторые гормоны, в том числе норадреналин и эпидермальный фактор роста (EGF), а также другие сигналы, такие как свет (с родопсином), вызывают фосфорилирование белка. EGF стимулирует рост нормальных фибробластов путем связывания со специфическим трансмембранным белковым рецептором на поверхности клетки. Гормональный сигнал в этом случае передается путем самофосфорилирования рецептора на внутриклеточной стороне после связывания гормона с последующим катализируемым другими киназами фосфорилированием белков, интернализацией комплекса EGF-EGF рецептор и сложным набором последствий. в ядре и в других местах при подготовке к клеточному делению. Бремер и др. (1986) недавно обнаружили, что ганглиозид GM3 ингибирует этот процесс аллостерическим образом, предотвращая самофосфорилирование рецептора EGF после связывания EGF. Для этого GM3 во внешней половине клеточной мембраны должен взаимодействовать с доменом полипептидной цепи рецептора EGF, вероятно, вызывая конформационные изменения, препятствующие фосфорилированию.
Некоторые гормоны, в том числе норадреналин и эпидермальный фактор роста (EGF), а также другие сигналы, такие как свет (с родопсином), вызывают фосфорилирование белка. EGF стимулирует рост нормальных фибробластов путем связывания со специфическим трансмембранным белковым рецептором на поверхности клетки. Гормональный сигнал в этом случае передается путем самофосфорилирования рецептора на внутриклеточной стороне после связывания гормона с последующим катализируемым другими киназами фосфорилированием белков, интернализацией комплекса EGF-EGF рецептор и сложным набором последствий. в ядре и в других местах при подготовке к клеточному делению. Бремер и др. (1986) недавно обнаружили, что ганглиозид GM3 ингибирует этот процесс аллостерическим образом, предотвращая самофосфорилирование рецептора EGF после связывания EGF. Для этого GM3 во внешней половине клеточной мембраны должен взаимодействовать с доменом полипептидной цепи рецептора EGF, вероятно, вызывая конформационные изменения, препятствующие фосфорилированию. Аналогичная ситуация, связанная с липидной мембраной, наблюдается с митохондриальным ферментом бета-гидроксимасляной дегидрогеназой, который каталитически активен только при включении в липидный бислой, состоящий из определенных фосфолипидов. Компьютерное математическое моделирование таких надмолекулярных структур будет необходимо для более глубокого понимания организации биологических материалов для выполнения сложных функций.
Аналогичная ситуация, связанная с липидной мембраной, наблюдается с митохондриальным ферментом бета-гидроксимасляной дегидрогеназой, который каталитически активен только при включении в липидный бислой, состоящий из определенных фосфолипидов. Компьютерное математическое моделирование таких надмолекулярных структур будет необходимо для более глубокого понимания организации биологических материалов для выполнения сложных функций.
Сноски
- 1
На самом деле правильнее говорить «один цистрон — один полипептид». Это также уже не является строго точным, поскольку более чем один ген может вносить вклад в первичную структуру белка, то есть в иммуноглобулины.
3.4 Клеточная мембрана — концепции биологии — 1-е канадское издание
Перейти к содержимому
Глава 3. Введение в структуру и функции клетки
К концу этого раздела вы сможете:
- Понимать жидкостно-мозаичную модель мембран
- Опишите функции фосфолипидов, белков и углеводов в мембранах
Плазматическая мембрана клетки определяет границы клетки и определяет характер ее контакта с окружающей средой. Клетки исключают одни вещества, поглощают другие и выделяют третьи, и все это в контролируемых количествах. Плазматические мембраны охватывают границы клеток, но они не являются статичным мешком, они динамичны и постоянно находятся в движении. Плазматическая мембрана должна быть достаточно гибкой, чтобы позволить определенным клеткам, таким как эритроциты и лейкоциты, изменять форму при прохождении через узкие капилляры. Это наиболее очевидные функции плазматической мембраны. Кроме того, поверхность плазматической мембраны несет маркеры, которые позволяют клеткам распознавать друг друга, что жизненно важно, поскольку ткани и органы формируются во время раннего развития, и которые позже играют роль в различии «своих» и «чужих» клеток. иммунный ответ.
Клетки исключают одни вещества, поглощают другие и выделяют третьи, и все это в контролируемых количествах. Плазматические мембраны охватывают границы клеток, но они не являются статичным мешком, они динамичны и постоянно находятся в движении. Плазматическая мембрана должна быть достаточно гибкой, чтобы позволить определенным клеткам, таким как эритроциты и лейкоциты, изменять форму при прохождении через узкие капилляры. Это наиболее очевидные функции плазматической мембраны. Кроме того, поверхность плазматической мембраны несет маркеры, которые позволяют клеткам распознавать друг друга, что жизненно важно, поскольку ткани и органы формируются во время раннего развития, и которые позже играют роль в различии «своих» и «чужих» клеток. иммунный ответ.
Плазматическая мембрана также несет рецепторы, являющиеся местами прикрепления специфических веществ, взаимодействующих с клеткой. Каждый рецептор устроен так, чтобы связываться с определенным веществом. Например, поверхностные рецепторы мембран создают изменения внутри, такие как изменения ферментов метаболических путей.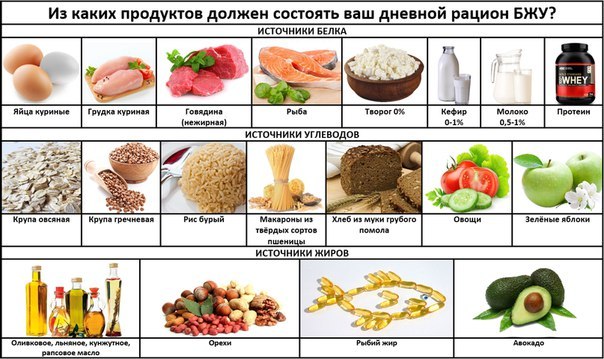 Эти метаболические пути могут иметь жизненно важное значение для обеспечения клетки энергией, производства специфических веществ для клетки или расщепления клеточных отходов или токсинов для утилизации. Рецепторы на внешней поверхности плазматической мембраны взаимодействуют с гормонами или нейротрансмиттерами и позволяют передавать их сообщения в клетку. Некоторые сайты распознавания используются вирусами в качестве точек присоединения. Хотя они очень специфичны, такие патогены, как вирусы, могут эволюционировать, чтобы использовать рецепторы для проникновения в клетку, имитируя конкретное вещество, которое рецептор должен связывать. Эта специфичность помогает объяснить, почему вирус иммунодефицита человека (ВИЧ) или любой из пяти типов вирусов гепатита проникают только в определенные клетки.
Эти метаболические пути могут иметь жизненно важное значение для обеспечения клетки энергией, производства специфических веществ для клетки или расщепления клеточных отходов или токсинов для утилизации. Рецепторы на внешней поверхности плазматической мембраны взаимодействуют с гормонами или нейротрансмиттерами и позволяют передавать их сообщения в клетку. Некоторые сайты распознавания используются вирусами в качестве точек присоединения. Хотя они очень специфичны, такие патогены, как вирусы, могут эволюционировать, чтобы использовать рецепторы для проникновения в клетку, имитируя конкретное вещество, которое рецептор должен связывать. Эта специфичность помогает объяснить, почему вирус иммунодефицита человека (ВИЧ) или любой из пяти типов вирусов гепатита проникают только в определенные клетки.
В 1972 году С. Дж. Сингер и Гарт Л. Николсон предложили новую модель плазматической мембраны, которая, по сравнению с более ранним пониманием, лучше объясняла как микроскопические наблюдения, так и функцию плазматической мембраны. Это было названо жидкостно-мозаичной моделью. Модель несколько эволюционировала с течением времени, но по-прежнему лучше всего описывает структуру и функции плазматической мембраны, как мы их сейчас понимаем. Жидкостно-мозаичная модель описывает структуру плазматической мембраны как мозаику компонентов, включая фосфолипиды, холестерин, белки и углеводы, в которой компоненты могут течь и менять положение 9.0114, при сохранении основной целостности мембраны. Как молекулы фосфолипидов, так и встроенные белки способны быстро и латерально диффундировать в мембрану. Текучесть плазматической мембраны необходима для деятельности определенных ферментов и транспортных молекул внутри мембраны. Плазматические мембраны имеют толщину от 5 до 10 нм. Для сравнения, эритроциты человека, видимые с помощью световой микроскопии, имеют толщину примерно 8 мкм, или примерно в 1000 раз толще плазматической мембраны.
Это было названо жидкостно-мозаичной моделью. Модель несколько эволюционировала с течением времени, но по-прежнему лучше всего описывает структуру и функции плазматической мембраны, как мы их сейчас понимаем. Жидкостно-мозаичная модель описывает структуру плазматической мембраны как мозаику компонентов, включая фосфолипиды, холестерин, белки и углеводы, в которой компоненты могут течь и менять положение 9.0114, при сохранении основной целостности мембраны. Как молекулы фосфолипидов, так и встроенные белки способны быстро и латерально диффундировать в мембрану. Текучесть плазматической мембраны необходима для деятельности определенных ферментов и транспортных молекул внутри мембраны. Плазматические мембраны имеют толщину от 5 до 10 нм. Для сравнения, эритроциты человека, видимые с помощью световой микроскопии, имеют толщину примерно 8 мкм, или примерно в 1000 раз толще плазматической мембраны.
Рис. 3.21 Жидкостно-мозаичная модель структуры плазматической мембраны описывает плазматическую мембрану как жидкую комбинацию фосфолипидов, холестерина, белков и углеводов.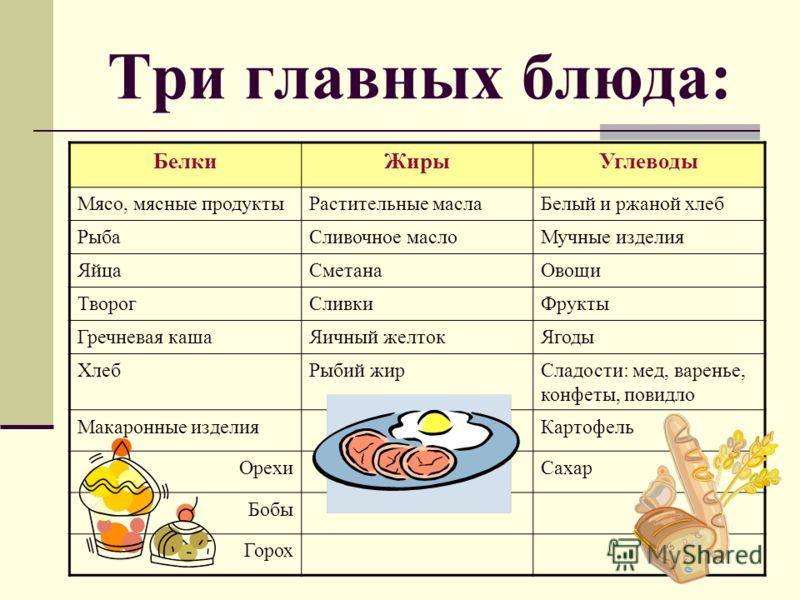
Плазматическая мембрана состоит в основном из двойного слоя фосфолипидов со встроенными белками, углеводами, гликолипидами и гликопротеинами, а в клетках животных — холестерином. Количество холестерина в плазматических мембранах животных регулирует текучесть мембраны и изменяется в зависимости от температуры окружающей среды клетки. Другими словами, холестерин действует как антифриз в клеточной мембране и более распространен у животных, живущих в холодном климате.
Основная ткань мембраны состоит из двух слоев молекул фосфолипидов, причем полярные концы этих молекул (выглядящие как набор шариков в художественном исполнении модели) (рис. 3.22) контактируют с водной жидкостью как внутри и снаружи клетки. Таким образом, обе поверхности плазматической мембраны гидрофильны. Напротив, внутренняя часть мембраны между двумя ее поверхностями представляет собой гидрофобную или неполярную область из-за хвостов жирных кислот. Эта область не имеет притяжения для воды или других полярных молекул.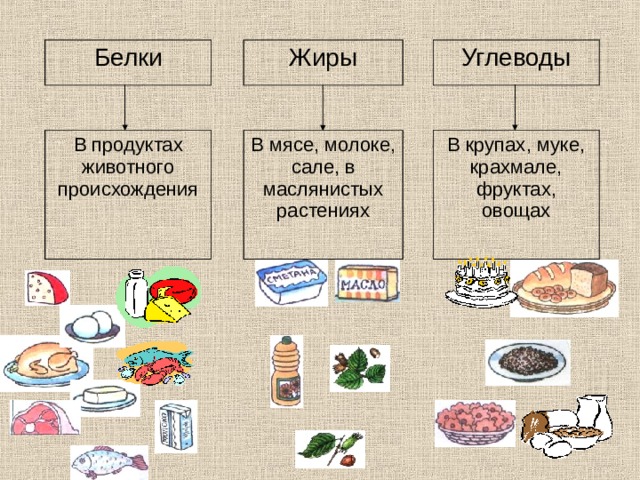
Рис. 3.22. Эта молекула фосфолипида состоит из гидрофильной головки и двух гидрофобных хвостов. Гидрофильная головная группа состоит из фосфатсодержащей группы, присоединенной к молекуле глицерина. Гидрофобные хвосты, содержащие либо насыщенную, либо ненасыщенную жирную кислоту, представляют собой длинные углеводородные цепи.
Белки составляют второй основной химический компонент плазматических мембран. Интегральные белки встроены в плазматическую мембрану и могут охватывать всю или часть мембраны. Интегральные белки могут служить каналами или насосами для перемещения материалов в клетку или из нее. Периферийные белки находятся на внешней или внутренней поверхности мембран, прикрепленные либо к интегральным белкам, либо к молекулам фосфолипидов. Как интегральные, так и периферические белки могут служить ферментами, структурными прикреплениями к волокнам цитоскелета или частью клеточных сайтов узнавания.
Углеводы являются третьим основным компонентом плазматических мембран. Они всегда находятся на внешней поверхности клеток и связаны либо с белками (образуя гликопротеины), либо с липидами (образуя гликолипиды). Эти углеводные цепи могут состоять из 2–60 моносахаридных звеньев и могут быть прямыми или разветвленными. Наряду с периферическими белками углеводы образуют на клеточной поверхности специализированные участки, позволяющие клеткам узнавать друг друга.
Они всегда находятся на внешней поверхности клеток и связаны либо с белками (образуя гликопротеины), либо с липидами (образуя гликолипиды). Эти углеводные цепи могут состоять из 2–60 моносахаридных звеньев и могут быть прямыми или разветвленными. Наряду с периферическими белками углеводы образуют на клеточной поверхности специализированные участки, позволяющие клеткам узнавать друг друга.
Эволюция в действии
Как вирусы заражают определенные органы Специфические молекулы гликопротеина, экспонированные на поверхности клеточных мембран клеток-хозяев, используются многими вирусами для заражения определенных органов. Например, ВИЧ способен проникать через плазматические мембраны определенных видов лейкоцитов, называемых Т-хелперами и моноцитами, а также через некоторые клетки центральной нервной системы. Вирус гепатита атакует только клетки печени.
Эти вирусы способны вторгаться в эти клетки, потому что клетки имеют сайты связывания на своей поверхности, которые вирусы используют с одинаково специфическими гликопротеинами в своей оболочке.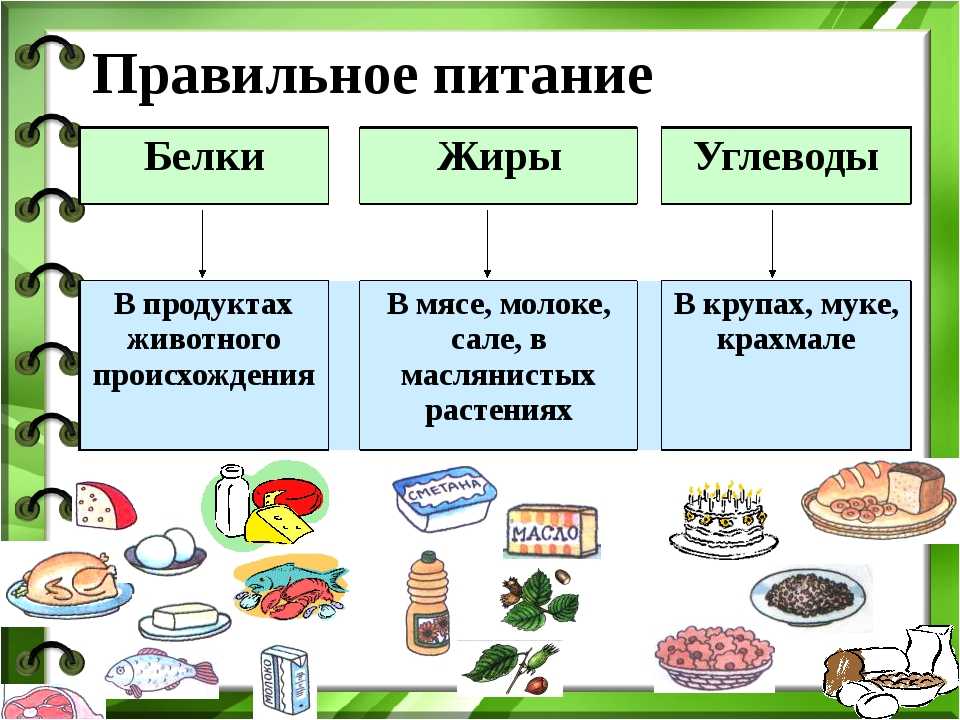 (рис. 3.23). Клетка обманывается мимикрией молекул оболочки вируса, и вирус может проникнуть в клетку. Другие участки распознавания на поверхности вируса взаимодействуют с иммунной системой человека, побуждая организм вырабатывать антитела. Антитела вырабатываются в ответ на антигены (или белки, связанные с инвазивными патогенами). Эти же участки служат местами для прикрепления антител и либо уничтожают, либо подавляют активность вируса. К сожалению, эти участки на ВИЧ кодируются генами, которые быстро меняются, что очень затрудняет производство эффективной вакцины против вируса. Популяция вируса внутри инфицированного человека быстро эволюционирует посредством мутаций в разные популяции или варианты, отличающиеся различиями в этих сайтах узнавания. Это быстрое изменение вирусных поверхностных маркеров снижает эффективность иммунной системы человека в борьбе с вирусом, поскольку антитела не распознают новые вариации поверхностных паттернов.
(рис. 3.23). Клетка обманывается мимикрией молекул оболочки вируса, и вирус может проникнуть в клетку. Другие участки распознавания на поверхности вируса взаимодействуют с иммунной системой человека, побуждая организм вырабатывать антитела. Антитела вырабатываются в ответ на антигены (или белки, связанные с инвазивными патогенами). Эти же участки служат местами для прикрепления антител и либо уничтожают, либо подавляют активность вируса. К сожалению, эти участки на ВИЧ кодируются генами, которые быстро меняются, что очень затрудняет производство эффективной вакцины против вируса. Популяция вируса внутри инфицированного человека быстро эволюционирует посредством мутаций в разные популяции или варианты, отличающиеся различиями в этих сайтах узнавания. Это быстрое изменение вирусных поверхностных маркеров снижает эффективность иммунной системы человека в борьбе с вирусом, поскольку антитела не распознают новые вариации поверхностных паттернов.
Рис. 3.23. ВИЧ стыкуется и связывается с рецептором CD4, гликопротеином на поверхности Т-клеток, прежде чем проникнуть в клетку или заразить ее.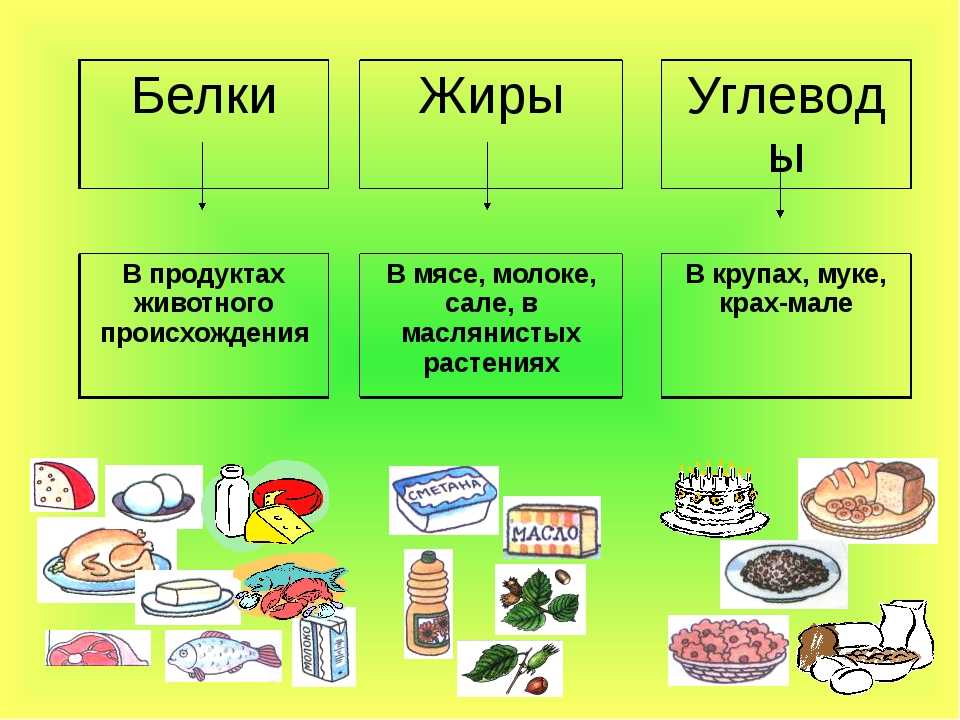
Современное понимание плазматической мембраны называется жидкостно-мозаичной моделью. Плазматическая мембрана состоит из двойного слоя фосфолипидов, гидрофобные жирнокислотные хвосты которых контактируют друг с другом. Ландшафт мембраны усеян белками, некоторые из которых пересекают мембрану. Некоторые из этих белков служат для транспортировки материалов в клетку или из нее. Углеводы присоединены к некоторым белкам и липидам на внешней поверхности мембраны. Они образуют комплексы, которые функционируют для идентификации клетки с другими клетками. Жидкостная природа мембраны обусловлена конфигурацией хвостов жирных кислот, наличием встроенного в мембрану холестерина (в клетках животных) и мозаичностью белков и белково-углеводных комплексов, не закрепленных прочно в место. Плазматические мембраны охватывают границы клеток, но они не являются статичным мешком, они динамичны и постоянно находятся в движении.
жидкостно-мозаичная модель: модель структуры плазматической мембраны в виде мозаики компонентов, включая фосфолипиды, холестерин, белки и гликолипиды, что приводит к жидкостному, а не статическому характеру
Атрибуция СМИ
- Рисунок 3.