Физиологическая роль мышечной системы: Физиология мышечной системы организма
Физиология мышечной системы организма
Мышечная система
Опорно-двигательный аппарат организма состоит примерно из 700 мышц, из них 400 скелетные мышцы, привлекаемые организмом для производства движения.
Мышечные волокна активируются командами нейронов ЦНС (нервно-мышечный синапс). Нерв и мышечные волокна, которые он активирует, называются моторная бляшка (моторная единица, двигательная единица).
Моторная бляшка (двигательная единица) работает по принципу все или ничего «All or None». Когда нерв получает сигнал от ЦНС на выполнение сокращения, это приводит к одновременному сокращению всех мышечных волокон, соединенных с одной моторной бляшкой. В то время, как без команды не происходит ровным счетом ничего.
Мышечные волокна делятся на две основные группы:
Красные волокна, медленные (тип I, ST — Slow Twitch), с высокой аэробной способностью.
Белые волокна, быстрые (тип II, FT — Fast Twitch), с высоким потенциалом для анаэробных упражнений.
Красные и белые волокна отличаются по своим физиологическим и анатомическим особенностям, в соответствии с их различными возможностями.
Рост мышечной силы напрямую зависит от взаимодействия и координации пары нерв — мышца (привлечение моторных бляшек), а они в свою очередь от процесса гипертрофии, в котором происходит рост объема мышечных волокон. Гиперплазия процесс, в котором происходит рост количества мышечных клеток, не может осуществляться в скелетных мышцах человека.
Два основных защитных механизма мышц:
• Мышечное веретено (Muscle Spindle), чье основное предназначение информировать ЦНС о длине мышечного волокна и влиять на величину сокращения и укорачивания мышцы, предотвращая перенапряжение.
• Сухожильный орган Гольджи (Golgi Tendon Organ), передает информацию нервной системе, о силе нагрузке на мышцы, и тем самым при превышении порога включает механизм расслабления и удлинения мышечных волокон.
Непривычная нагрузка на мышцы приводит в последствии к болевым ощущениям, которые появляются через 24-48 часов с момента окончания мышечной работы. «Поздняя мышечная боль» — (DOMS — Delayed Onset Muscle Soreness) выражается в уменьшении силы мышц и ощущении болезненной чувствительности, жесткости и боли в мышцах . DOMS не требует медицинского лечения, но рекомендуется сочетать активное восстановление (упражнения низкой интенсивности) вместе с пассивными способами (массаж, сауна, горячая ванна). По истечении примерно 5 дней боли проходят, мышца восстанавливается и готова к нагрузкам очередной тренировки.
По истечении примерно 5 дней боли проходят, мышца восстанавливается и готова к нагрузкам очередной тренировки.
Внимание! У Вас нет прав для просмотра скрытого текста.
Ключевые теги: физиология мышц
Физиология нервно-мышечной системы. — Атшибаева Оксана
Скелетные мышцы состоят из мышечных клеток, называемых волокнами. Волокна соединены в пучок и покрыты оболочкой из соединительной ткани. В состав каждой мышцы входит от ста до десятков тысяч волокон.
Диаметр мышечного волокна от 12 до 70 мкм, а его длина 10-12 см.
Каждое мышечное волокно окружено тонкой эластичной оболочкой – сарколеммой. Внутри волокна находиться саркоплазма. Длинные нитевидные образования, тянущиеся от одного конца волокна к другому, наз. мифибриллами. Каждая мифибрилла состоит еще с более тонких нитей – протифибрилл. Толстые протифибриллы состоят из белка миозина, а тонкие – из белка актина. Они расположены в строгом порядке по всей длине мифибриллы, что предает волокну поперечно-полосатую исчерченность. Поэтому скелетные мышцы и называются поперечно-полосатые. Нити миозина и актина являются сократительным аппаратом мышечного волокна.
Поэтому скелетные мышцы и называются поперечно-полосатые. Нити миозина и актина являются сократительным аппаратом мышечного волокна.
строение нервно-мышечного волокна
В состав мышечной клетки входят митохондрии. Они играют важную роль в окислительных процессах и энергетическом обмене. В мышечном волокне много ядер.
функции митохондрии
Мышечное волокно характеризируется такими физиологическими свойствами:
- Возбудимостью, т.е. способностью отвечать возбуждением на раздражение.
- Проводимостью, т.е. способностью проводить возбуждение в обе стороны от места раздражения по всей длине.
- Сократимостью, т.е. способностью сокращаться или изменять степень напряжения при возбуждении.
Скелетные мышцы – активная часть двигательного аппарата.
Их функции:
- Перемещать тело в пространстве (ходьба, бег, плавание)
- Перемещают части тела относительно друг друга (движение конечностей, головы, туловища и т.
 д.)
д.) - Поддерживают положение тела в определенной позе (сидячей, стоячей и т.д.)
 д.)
д.)
Функция нервного волокна состоит в проведении импульсов, что подчинено определенным законам:
- Закон двустороннего проведения возбуждения: возбуждение распространяется по нервному волокну в обе стороны от места нанесения раздражения.
- Закон изолированного проведения возбуждения: возбуждение распространяется только по одному нервному волокну, не переходя на другие.
- Закон физиологической непрерывности: для проведения возбуждения необходима структурная и функциональная целостность нервного волокна. Нервного волокно практически неутомляемо; его работоспособность по сравнению с другими возбудимыми тканями значительно выше.
нервное волокно в мышце
Основная функция мышцы – сокращение, т.е. изменение напряжения. Для этого необходимо, чтобы возбуждение перешло с нерва на мышцу.
Возбуждение с нерва на мышцу передается с помощью особого химического вещества – медиатора. В нервно-мышечном соединении медиатором служит ацетилхолин. Он образуется в организме из витамина холина, относящегося к витаминам группы В. Под влиянием ацетилхолина изменяется электрический заряд мышечного волокна, вызывает возбуждение.
В нервно-мышечном соединении медиатором служит ацетилхолин. Он образуется в организме из витамина холина, относящегося к витаминам группы В. Под влиянием ацетилхолина изменяется электрический заряд мышечного волокна, вызывает возбуждение.
Сокращение мышечного волокна происходит за счет энергии, источником является аденозинтрифосфорная кислота (АТФ).
Трудовые процессы требуют чрезвычайно тонкой регуляции мышечного напряжения.
Во первых, мышечное напряжение зависит от количества активных, работающих, двигательных единиц (нервная клетка, нерв, нервно-мышечное соединение, мышечное волокно). В состав одной мышцы входит несколько тысяч двигательных единиц. Обычно только часть из них находится в активном состоянии. Чем больше активных двигательных единиц, тем больше напряжение мышцы.
Во-вторых, мышечное напряжение зависит частоты импульсации управляющего ею мотонейрона: чем выше частота импульса, тем больше активность двигательной единицы и тем больше напряжение возникает в ее мышечных волокнах.
В-третьих, в регуляции мышечного напряжения играет роль согласование активности разных двигательных единиц во времени. Двигательные единицы сокращаются одновременно (синхронно) и поочередно (асинхронно). Синхронное сокращение группы двигательных единиц обеспечивает большое напряжение, чем асинхронное. При асинхронной работе двигательных мышц дольше поддерживается напряжение, движения совершаются более плавно. В обычных условиях большинство двигательных единиц работает асинхронно. При утомлении они начинают работать синхронно. Внешне это проявляется в нарушении плавности и точности движений, в возникновании дрожания (тремора).
Физиологическая роль мышц — презентация онлайн
2. Физиологическая роль мышц
1.
2.
3.
4.
5.
6.
7.
Поддержание позы и равновесия тела
Произвольные движения
Воспроизведение речи
Работа сердца
Регуляция тонуса сосудов
Моторика желудка и кишечника
Терморегуляция (сократительный
термогенез)
3.
 У детей
У детей
В постнатальном периоде до реализации
позы, функция мышечной системы в
значительной степени связана с
терморегуляцией. Для детей этого периода
характерна постоянная активность скелетной
мускулатуры. Постоянная активность мышц
является стимулом бурного роста мышечной
массы и скелета, правильного формирования
суставов.
В период реализации позы
терморегуляторная функция скелетной
мускулатры снижается, и тоническая форма
деятельности сменяется фазно-тонической.
4. Двигательная единица
Комплекс,
включающий один
мотонейрон и
иннервируемые им
мышечные волокна,
называется
ДВИГАТЕЛЬНОЙ
ИЛИ
НЕЙРОМОТОРНОЙ
ЕДИНИЦЕЙ
5. Два варианта классификации скелетных мышц
1. Анатомическая
По плотности иннервации
(количество двигательных
единиц в мышце)
2. Функциональная
По работоспособности
двигательных единиц
6. Классификация по количеству двигательных единиц в мышце
Много двигательных единиц на одну мышцу
Плотность иннервации высокая
1 мотонейрон иннервирует 10 – 25 волокон
В мышцах, приспособленных для тонких
движений (пальцы, язык, наружные мышцы
глаза).
Мало двигательных единиц на одну мышцу
Плотность иннервации низкая
1 мотонейрон иннервирует 700 – 1000 волокон)
В мышцах поддерживающих позу,
осуществляющих «грубые» движения
(мышцы туловища).
7. Классификация по работоспособности
1.Медленные
малоутомляемые
2.Быстрые
легкоутомляемые
Медленные
малоутомляемые
Быстрые
легкоутомляемые
Мотонейроны
Частота
импульсов
Сила,
сокращения
Количество
митохондрий
низкопороговые
До 10 Гц
высокопороговые
50-60 Гц
меньше
больше
Много
Мало
Запасы гликогена
Количество
миоглобина
Много
Много
Мало
Мало
Плотность
капилляров
Зависимость от
поступления О2
Много
Мало
Малочувствительны к Высокочувствитель
недостатку
ны к недостатку
9. Свойства мышцы
Возбудимость
Проводимость
Сократимость
Эластичность – способность
сокращаться при растягивании.
5. Тонус
1.
2.
3.
4.
10. Мышечное волокно
11. Саркомер
12. Возбуждение мышечных волокон
13. Одно мышечное волокно получает нервный импульс от одного синапса
14. Нервно-мышечный синапс (концевая пластинка)
15. Медиатор – ацетилхолин На постсинаптической мембране Н-холинорецепторы
16. Потенциал концевой пластинки
17. Особенности нервно-мышечного синапса
1. Большая поверхность
пресинаптической мембраны
2. Синаптическая щель содержит
много ГАГ, митохондрий
3. Большая складчатость
постсинаптической мембраны
4. Нет суммации – ПКП сразу
переходит в ПД
18. У детей
постсинаптическая мембрана не
сформирована, мышечное волокно
чувствительно к ацетилхолину на всем его
протяжении, а не только в непосредственной
близости нервного волокна. Постепенно
участок мышечной клетки, реагирующий на
ацетилхолин, становится меньше и
ограничивается небольшим участком,
соответствующим зоне синапса. У человека
У человека
расширение нервно-мышечного синапса,
усложнение его структуры (складчатость
постсинаптической мембраны, количество
медиатора, митохондрий) продолжается до
18 – 20 лет.
19. ПД распространяется по мембране мышечного волокна
20. Результат возбуждения — увеличение концентрации внутриклеточного кальция
21. Потенциал концевой пластинки (ПКП) и потенциал действия (ПД) мышечного волокна
Синапс
ПД
ПД
ПКП
Са++
СПР
22. У детей
Возбудимость мышц плода и детей низка,
т.к. мембранный потенциал покоя
значительно менее отрицателен, чем у
взрослых, примерно -20 — -40мВ, Это
объясняется небольшим количеством и
малой активностью ионных насосов,
недостаточным количеством ионных каналов
на мембране мышечных волокон.
Эти же причины лежат в основе низкой
проводимости мышечных волокон
новорожденных и детей. По мере взросления
скорость проведения ПД увеличивается и за
счет появления новых ионных каналов, и за
счет увеличения толщины мышечного
волокна.
23. Последовательность событий при возбуждении
1. ПД пресинаптического окончания приводит к
2.
3.
4.
5.
6.
выделению медиатора.
Возникновение ПКП на постсинаптической
мембране.
Возникновение ПД — возбуждение мембраны.
Проведение возбуждения по Т-системе
высвобождение Ca++ из
саркоплазматического ретикулума (СПР).
Концентрация ионов увеличивается с с 10-8
до 10-5 моль.
Взаимодействие Са++ с тропонином
24. Сокращение
Сократительные и
регуляторные белки
саркомера
25. Саркомер – функциональная единица сократительного аппарата мышечной клетки. Длина саркомера 2,5 мкм, поперечник – 1мкм
Саркомер
–
функциональная
единица
сократительного аппарата мышечной
.
клетки.
Длина саркомера 2,5 мкм, поперечник – 1мкм
28. Толстые миофиламенты
29. Свойства миозина
Толстые миофиламенты – образованы молекулами
миозина, Тяжелые цепи миозина — головка миозина
и шейка.
Головка обладает АТФ-азной
активностью
Шейка обладает эластическими
свойствами.
В толстой филаменте 150 молекул миозина.
Под электронным микроскопом на толстой
миофиламенте видны выступы, расположенные
под углом 120 градусов. Они получили название
поперечных мостиков.
Эти мостики образованы головкой и шейкой молекул
миозина, их длина 20 нм.
30. Тонкие миофиламенты
31. Свойства актина
Тонкие миофиламенты построены из
глобулярных молекул белка актина.
Актиновые филаменты представляют
собой закрученную двойную спираль
Таких нитей в саркомере 2000.
Они одним концом прикреплены к пластинке, а второй конец достигает
середины саркомера.
32. Регуляторные белки
В продольных бороздках актиновой
спирали располагаются
нитевидные молекулы белка
тропомиозина. Тропомиозин
закрывает активные центры
на актиновых нитях
К молекуле тропомиозина
равномерно прикреплены
молекулы тропонина. Этот белок
Этот белок
может связывать катионы Са++
34. Контакт актина с миозином возможен, если тропомиозин сдвинется и откроет активные центры на тонких – актиновых — миофиламентах
35. Ключевой момент – переход отвозбуждения к сокращению
Связывание ионов кальция
молекулами тропонина
Изменение конформации
тропомиозина
Сдвиг тропомиозина
Освобождение активных центров
актиновых миофиламентов
37. «гребок»
В момент контакта головка миозина
совершает «гребковое» движение и
передвигает тонкую миофиламенту
вдоль толстой по направлению к центру
саркомера
38. Укорочение саркомера
45. Последовательность событий при сокращении.
1. молекулы
2.
3.
4.
5.
тропомиозина опускаются в
желобки между цепочками
мономеров
актина, открывая участки прикрепления для
поперечных мостиков миозина.
Электромеханическое сопряжение
Прикрепление головки миозина к активному
центру в актиновых нитях и повышение АТФазной активности.
Гребковое движение шейки и перемещение
нитей актина относительно миозина.
Поворот головки и увеличение напряжения
шейки
Гидролиз АТФ, отрыв и прикрепление к
следующему активному центру
46. Расслабление
Работа Са++ АТФ-азы
Снижение концентрации
внутриклеточного Са++
Восстановление конформации
тропомиозина
Активные центры тонких
миофиламентов закрыты.
47. Временная характеристика процесса сокращения
48. Сопоставление во времени сокращения мышцы и возбудимости
Возбудимость
мышцы во время
сокращения
нормальна,
следовательно
она может
ответить на
раздражение
49. Запись мышечных сокращений
50. Суммация сокращений
51. Режимы мышечного сокращения
1. Одиночное сокращение
2. Тетанус – слитное сокращение без
расслабления
a) Зубчатый – импульс в фазу
расслабления
b) Гладкий – импульс в фазу укорочения
3. Оптимум – частота для включения всех
двигательных единиц
4. Пессимум – частота за пределами
Пессимум – частота за пределами
функциональной лабильности
52. Режимы сокращения
1. Одиночное
2. Зубчатый
тетанус
3. Гладкий тетанус
4. Оптимум
СКЕЛЕТНЫЕ МЫШЦЫ
НОВОРОЖДЕННЫХ НЕ СПОСОБНЫ
РАЗВИВАТЬ ТЕТАНУС — низкая
функциональная лабильность ( высокая
продолжительность ПД).
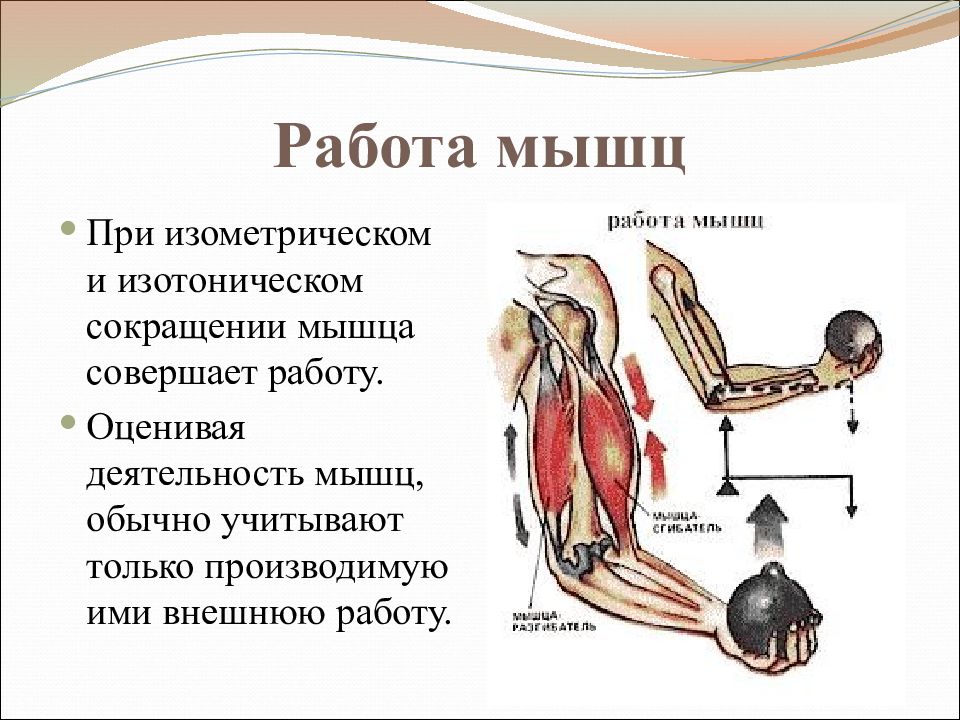
54. Виды мышечного сокращения
Виды
1. Изометрическое
2. Изотоническое
3. Ауксотоническое
55. Регуляция силы сокращения мышцы
Сила сокращения зависит от
1. числа включенных мышечных
волокон
2. частоты импульсов возбуждения
– режима сокращения
3. степени синхронизации частот
для всех волокон
56. Законы мышечного сокращения
1. Мышечное волокно подчиняется
закону все или ничего
2. Целая мышца подчиняется закону
силы (число включенных
мышечных волокон, режим их
работы)
Состав питьевой воды от «Родник Прикамья»
Кальций
- строительный материал для роста костей, зубов, волос, ногтей
- нормализует кислотно-щелочное равновесие в организме человека
- оказывает противовоспалительное и противоаллергическое действие
- участвует в механизме свертывания крови
- важен для нормальной работы мышечной системы
- регулирует нормальный ритм сердечной мышцы
- участвует в проведении нервных импульсов.

Магний
- участвует в углеводном и фосфорном обмене
- обладает сосудорасширяющим и противоспастическим действием
- стимулирует перистальтику кишечника и повышает желчеотделение
- магний, как и кальций, играет важную роль в образовании и поддержании костной ткани.
Калий
- помогает регулировать водный и солевой баланс, обеспечивая постоянство внутренней среды в организме
- участвует в обмене белков и углеводов, необходим для их синтеза
- входит в состав транспортных систем клеточной мембраны, является компонентом калий-натриевого насоса
- обеспечивает нормальное функционирование нервных клеток, участвует в передаче нервного импульса
- необходим для нормальной работы мышц, в т. ч. сердечной
- выводит излишки воды и натрия из организма.
Фосфор
- вместе с кальцием является основным компонентом костной и зубной ткани
- активно участвует в обмене белков, жиров и углеводов
- необходим для передачи наследственной информации (входит в состав ДНК и РНК)
- участвует в энергообеспечении организма (входит в АТФ)
- необходим для регуляции рН (кислотно-щелочного баланса) внутренних жидких сред
Бикарбонаты
- Помогают поддерживать постоянный уровень кислотно-щелочного баланса в организме.

Роль физиологически необходимых микроэлементов:
Йод
- Поддерживает нормальную деятельность щитовидной железы
- Участвует в построении гормона тироксина щитовидной железой.
Физиологическая роль тироксина очень велика:
- контролирует интенсивность основного обмена веществ в организме
- оказывает влияние на водно-солевой, жировой и углеводный обмены
Гормон щитовидной железы находится в постоянной связи с другими железами внутренней секреции, особенно с гипофизом и половыми железами.
- активно воздействует на физическое и психическое развитие человека, влияет на эмоциональный тонус человека
- участвует в регуляции деятельности центральной нервной системы
- оказывает влияние на деятельность сердечно-сосудистой системы и печени человека.
Фтор
- участвует в костеобразовании и процессах формирования дентина и зубной эмали
- стимулирует кроветворную систему и иммунитет
- участвует в развитии скелета
- стимулирует восстановительные процессы при переломах костей.

Недостаток поступления фтора в организм является одним из факторов кариеса зубов, особенно в период их прорезывания и минерализации.
Общая минерализация воды
Вода высшей категории «Аква Лайт» имеет физиологически полноценную минерализацию (сумма всех растворимых в воде веществ — ионов, биологически активных элементов). Общая потребность организма человека в воде — 1,5-2 л в день.
Вода, имеющая показатели общей минерализации свыше 1000 мг./л, — является минеральной, пить такую воду можно с соблюдением рекомендуемого количества, а лечебные минеральные воды вообще рекомендуется пить только по назначению врача. Воду высшей категории качества «Аква Лайт» можно пить для удовлетворения полной потребности человека в питьевой воде.
Водородный показатель PH (кислотно-щелочной показатель)
Организм человека поддерживает постоянство внутренней среды, кислотно щелочной показатель основных биологических жидкостей имеет нейтральное значение pH 7,0-7,5, сдвиг равновесия в сторону закисления ведет к развитию заболеваний.
pH воды высшей категории «Аква Лайт» — 7,6, что соответствует физиологической норме и благоприятно для здоровья человека.
Жесткость воды (содержание растворимых солей кальция и магния)
Рекомендованный показатель общей жесткости для питьевой воды высшей категории качества (ГОСТ 4151) составляет 1,5-7,0 мг-экв/л. Вода «Аква Лайт» компании «Родник Прикамья» имеет показатель жесткости 3,1 мг-экв/л.
Устойчивой тенденцией последних лет является стремление людей к здоровому образу жизни. И качественная питьевая вода стоит в списке ценностей для здоровья человека на одном из первых мест. Компания «Родник Прикамья» предлагает питьевую воду высшей категории качества «Аква Лайт». Пейте и будьте здоровы.
Заказать воду высшей категории качества «Аква лайт» | Скачать прайс
Физиология мышечной и нервной систем: Учебное пособие. 1-е изд
Описание товара
Учебное пособие посвящено важному разделу физиологии, необходимому для теоретической подготовки и практической деятельности различных специалистов. В доступной форме в нем изложены основные закономерности функционирования мышечной и нервной систем, что позволяет изучить основные механизмы и закономерности их регуляции в живом организме. Благодаря емкому раскрытию материала учебное пособие удобно при подготовке к экзамену.
В доступной форме в нем изложены основные закономерности функционирования мышечной и нервной систем, что позволяет изучить основные механизмы и закономерности их регуляции в живом организме. Благодаря емкому раскрытию материала учебное пособие удобно при подготовке к экзамену.
Книга предназначена для студентов вузов, обучающихся по направлениям подготовки (специальностям) «Ветеринария», «Зоотехния», «Биология», а также будет полезна слушателям факультетов постдипломного образования, преподавателям вузов и научным работникам
Учебное пособие «Физиология мышечной и нервной систем» — под общ. ред. профессора И. Н. Медведева
Осуществление одной клеткой разнообразных функций характерно для одноклеточных животных, у многоклеточных организмов существует специализация клеток — разделение их функций. Так, мышечные клетки обладают способностью к сокращению, а нервные (нейроны) воспринимают информацию от внешних и внутренних раздражителей, перерабатывают ее и передают в виде сигналов на другие нейроны, внутренние органы и органы движения.
Примером обладателей наиболее простых форм нервной системы являются беспозвоночные животные, имеющие диффузную нервную сеть (медузы, гидры, губки и актинии). Возбуждение по нейронам такой сети распространяется диффузно, во все стороны, благодаря многочисленным взаимным связям между нейронами.
У высших беспозвоночных животных (насекомых, червей) нервная система усложняется, становится сегментарной. Тело этих животных разделено на сегменты (одинаковые участки), которые иннервируются нервными клетками, находящимися в этих сегментах.
По мере развития у позвоночных головного конца нервной системы (энцефалона) — головного мозга и формирования в нем различных нервных центров — групп нервных клеток, управляющих отдельными функциями, усложняются поведенческие реакции животных. По мере усиления процесса кортикализации, кора больших полушарий подчиняет себе все другие отделы нервной системы.
В ходе жизнедеятельности организма происходит его постоянное приспособление к окружающей среде или, наоборот, приспособление им среды к нему. Опираясь на весь накопленный современной наукой багаж знаний, живой организм можно охарактеризовать как саморегулирующуюся систему, являющуюся единым целым и адекватно реагирующую на все изменения окружающей среды. Это возможно благодаря тесной взаимосвязи и взаимоподчиненности процессов, происходящих в организме, а также взаимодействию между собой всех клеток и в первую очередь составляющих нервную и мышечную системы.
Опираясь на весь накопленный современной наукой багаж знаний, живой организм можно охарактеризовать как саморегулирующуюся систему, являющуюся единым целым и адекватно реагирующую на все изменения окружающей среды. Это возможно благодаря тесной взаимосвязи и взаимоподчиненности процессов, происходящих в организме, а также взаимодействию между собой всех клеток и в первую очередь составляющих нервную и мышечную системы.
Нервная система воспринимает внешние и внутренние раздражители, перерабатывает информацию и передает импульсацию на мышцы, стимулируя деятельность, иначе говоря, регулируя мышечный (двигательный) аппарат, благодаря работе которого происходит движение организма и его частей. Именно в результате адекватной двигательной деятельности организм активно реагирует на окружающую среду и приспосабливается к ней, выживает и оставляет потомство.
Оглавление
Введение ………. 5
Глава 1. Физиология центральной нервной системы ………. 7
1. 1. Общие вопросы функционирования центральной нервной системы ………. 7
1. Общие вопросы функционирования центральной нервной системы ………. 7
1.2. Общефункциональные особенности нейронов ………. 9
1.3. Физиологические основы процессов возбуждения в нервной клетке ………. 11
1.4. Синаптические связи между нейронами ………. 13
1.5. Пре- и постсинаптическое торможение ………. 18
1.6. Импульсация в нейронах. ………. 21
Контрольные вопросы к главе 1 ………. 25
Глава 2. Физиология сенсорных систем ………. 26
2.1. Строение и функции сенсорных систем ………. 26
2.2. Основы физиологии рецепторов ………. 27
2.3. Функционирование отдельных типов рецепторов ………. 31
Светочувствительные рецепторы зрительного анализатора ………. 31
Слуховые рецепторы ………. 35
Вестибулярные рецепторы ………. 38
Проприорецепторы двигательной сенсорной системы ………. 41
2.4. Общие вопросы физиологии сенсорных систем ………. 45
Процесс обработки афферентной информации . ……… 45
……… 45
Поля сенсорных систем в коре ………. 45
Пространственное представительство. ………. 47
Процессы активного восприятия ………. 48
Контрольные вопросы к главе 2 ………. 48
Глава 3. Физиология моторной системы ………. 49
3.1. Основные функции моторной системы ………. 49
3.2. Спинномозговые рефлексы двигательного характера ………. 50
Функционирование спинного мозга ………. 50
Рефлексы, возникающие на растяжение (миотатические рефлексы) ………. 53
Значение гамма-регуляции для мышечного тонуса ………. 54
Реципрокное функционирование центров мышц-антагонистов ………. 55
Сгибательные рефлексы ………. 56
Разгибательные рефлексы ………. 56
Особенности ритмических рефлексов ………. 57
3.3. Основные моторные центры головного мозга ………. 58
Моторные стволовые центры ………. 58
Функционирование подкорковых узлов. ….. ….. 60
….. 60
Мозжечок ………. 61
3.4. Двигательные функции коры больших полушарий 62
Вертикальные колонки нейронов в коре ………. 62
Роль отделов коры больших полушарий в обеспечении произвольных движений ………. 64
Электрические явления в коре больших полушарий 65
3.5. Медиальная и латеральная нисходящие моторные системы ………. 67
Пирамидальная корково-спинномозговая система ………. 68
Корково-красноядерно-спинномозговая система ………. 69
Медиальная система (вестибуло-спинномозговой и ретикулярно-спинномозговой компоненты) ………. 70
3.6. Основы регуляции двигательной деятельности ………. 71
Многоуровневая циклическая система регуляции движений ………. 71
Программирование, сенсорные коррекции и обратные связи ………. 73
Регуляция позы и движений ………. 75
Установочные рефлексы ………. 75
Роль отделов ЦНС в обеспечении позно-тонической мышечной активности . ……… 76
……… 76
Роль ЦНС в регуляции движений ………. 77
Контрольные вопросы к главе 3 ………. 80
Глава 4. Физиология высшей нервной деятельности ………. 81
4.1. Высшая нервная деятельность в обеспечении поведенческих реакций и приспособлении к внешней среде ………. 81
4.2. Условные и безусловные рефлексы ………. 82
4.3. Условия и механизмы, необходимые для образования условных рефлексов ………. 84
4.4. Условные рефлексы первого и второго рода ………. 87
4.5. Сложные и простые условные рефлексы ………. 88
4.6. Процесс торможения условных рефлексов ………. 94
4.7. Сигнальные системы ………. 97
4.8. Процессы экстраполяции в высшей нервной деятельности ………. 98
4.9. Типы высшей нервной деятельности ………. 99
4.10. Целенаправленное поведение ………. 101
4.11. Эмоции и память в формировании поведения ………. 103
Контрольные вопросы к главе 4 . ……… 104
……… 104
Глава 5. Физиология вегетативной нервной системы ………. 105
5.1. Симпатическая и парасимпатическая части вегетативной нервной системы ………. 106
5.2. Вегетативная иннервация ………. 110
Продолжительность эффекта ………. 110
Чувствительность регуляции ………. 110
Пусковые и настроечные воздействия ………. 111
Взаимодействие вегетативной и гуморальной регуляций ………. 112
5.3. Уровни рефлекторной регуляции вегетативных процессов ………. 113
Контрольные вопросы к главе 5 ………. 114
Глава 6. Физиология нервно-мышечного аппарата ………. 115
6.1. Морфология нервно-мышечного аппарата ………. 116
6.2. Проведение возбуждения с нерва на мышцу ………. 120
6.3. Механизм сокращения мышцы ………. 123
6.4. Энергетика и химизм сокращения мышц ………. 125
Фосфагенная энергетическая система ………. 127
Гликолитическая, или лактацидная, энергетическая система . ……… 128
……… 128
Окислительная система энергообеспечения ………. 132
6.5. Характеристики мышечного сокращения ………. 139
Формы и типы сокращения мышцы ………. 139
Понятие о характеристических кривых мышцы ………. 141
6.6. Режимы мышечного сокращения ………. 146
Одиночные сокращения. ………. 146
Тетаническое сокращение ………. 149
6.7. Быстрые и медленные двигательные единицы ………. 152
Мотонейроны: медленные и быстрые ………. 153
Быстрые и медленные мышечные волокна ………. 155
Состав мышц ………. 157
Подтипы быстрых мышечных волокон ………. 160
6.8. Регуляция мышечного напряжения ………. 161
Регуляция процесса напряжения мышцы ………. 162
Электромиограмма. ………. 166
Контрольные вопросы к главе 6 ………. 168
Заключение ………. 169
Список рекомендуемой литературы ………. 171
Лечение жевательной мышцы — лечение височно челюстного сустава
Обычно мы не задумываемся о том, как двигается наша челюсть.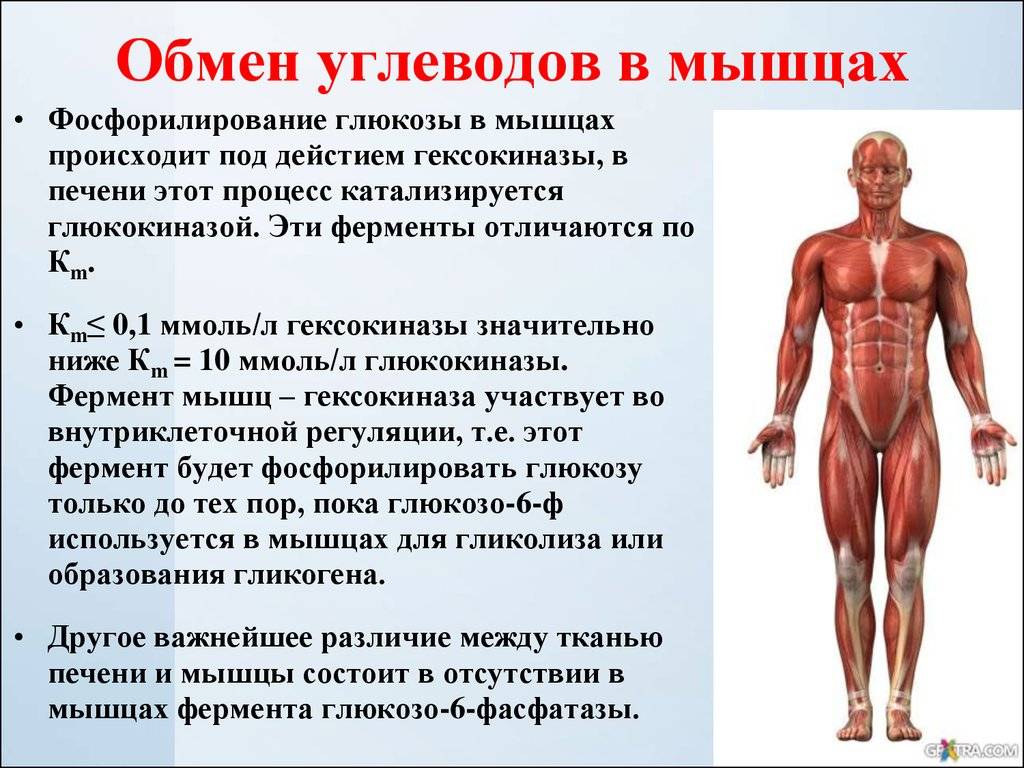 Большинство людей, говоря о прикусе, подразумевает зубы. Но, как упоминалось выше, прикус – это целая система, контролирующая положение челюсти. Вот ее составляющие и их роль:
Большинство людей, говоря о прикусе, подразумевает зубы. Но, как упоминалось выше, прикус – это целая система, контролирующая положение челюсти. Вот ее составляющие и их роль:
Роль мышц
Рисунок №4 Открывание, закрывание рта, жевание и глотание происходит благодаря работе большого количества мышц.
Открывание рта обеспечивают мышцы, располагающиеся в области шеи. Мышцы, удерживающие нижнюю челюсть, находятся позади челюсти и проходят под щеками к лобной и заушной областям.
Самые мощные мышцы, отвечающие за жевание, расположены в щечной области.
Рисунок №5 Наш организм стремится осуществлять все функции с наибольшей рациональностью и наименьшей затратой энергии. Поэтому в норме жевательные мышцы удерживают челюсть в положении, при котором зубы на 1-2 мм разобщены.
Рисунок №6 Из этого рационального положения нижняя челюсть сотни раз в день перемещается до смыкания зубов для осуществления глотания.
Все это происходит подсознательно. Наш мозг «запрограммирован» на процесс, который называется проприоцепция.
При правильном прикусе и положении челюсти в состоянии покоя большинство мышц находится в расслабленном состоянии.
Если Ваши зубы смыкаются не совсем правильно, мышцы могут приспосабливаться к этому, заставляя челюсть закрываться по траектории, вызывающей чрезмерное напряжение, спазм и со временем усталость мышц.
При этом положение физиологического покоя нижней челюсти (когда зубы не сомкнуты) не сопровождается расслаблением мышц, как это должно быть при нормальной окклюзии, а наоборот, приводит к их постоянному напряжению.
Роль суставов
Рисунок №7 Нижняя челюсть двигается благодаря суставам. называемым височнонижнечелюстными суставами, или ВНЧС.
Представьте сустав как мяч в корзине – суставная головка (мяч) двигается в углублении черепа (корзине). Между ними находится мягкотканная прослойка (суставной диск).
Положение суставной головки (мыщелка) в соответствующем углублении черепа (ямке) во многом определяется прикусом (окклюзией) – положением, где мышцы удерживают челюсть.
Рисунок №8 Если система сбалансирована, мыщелки расположены в центре суставной ямки и диск, находясь между суставной головкой и суставной ямкой, свободно двигается в суставе при открывании и закрывании рта.
Рисунок №9 Если жевательные мышцы приспосабливаются к неправильному положению челюсти, они могут удерживать головку сустава в смещенном положении, негативно влияя на ее функцию. Часто это приводит к смещению суставного диска, что препятствует нормальному его движению в суставе до того момента, пока он полностью не переместится в нормальное положение при открывании рта. Когда это происходит, возникает щелчок.
Интенсивность щелчка может варьироваться от едва осязаемой вибрации до очень громкого звука.
Этот звук часто является признаком ненормальной функции сустава, который в свою очередь, может быть связан с неправильным прикусом.
Роль осанки
Осанка также играет значительную роль в окклюзии. Нижнюю челюсть и нижние конечности можно считать взаимозависимыми частями скелетной системы человека. Если какая-либо из этих частей подвергается негативному влиянию, это может отразиться и на других частях скелета.
Проверим это, слегка сомкнув зубы. Обратите внимание на то, какие зубы сомкнулись первыми. Затем максимально откиньте голову назад и сомкните зубы еще раз. Не удивляйтесь, если на этот раз первый контакт придется на другие зубы. Изменяя положение головы, Вы тем самым влияете на баланс всей скелетной системы (и на Ваш прикус в частности).
То же самое происходит, когда Вам ставят пломбу. Лежа в стоматологическом кресле под анестезией кажется, что пломба не мешает. Но после того, как действие анестезии заканчивается, и Вы встаете с кресла, Вы замечаете, что пломба выше, чем остальные зубы. Вот почему в нейромышечной стоматологии прикус оценивают в положении сидя – положении, в котором обычно находится ваша челюсть.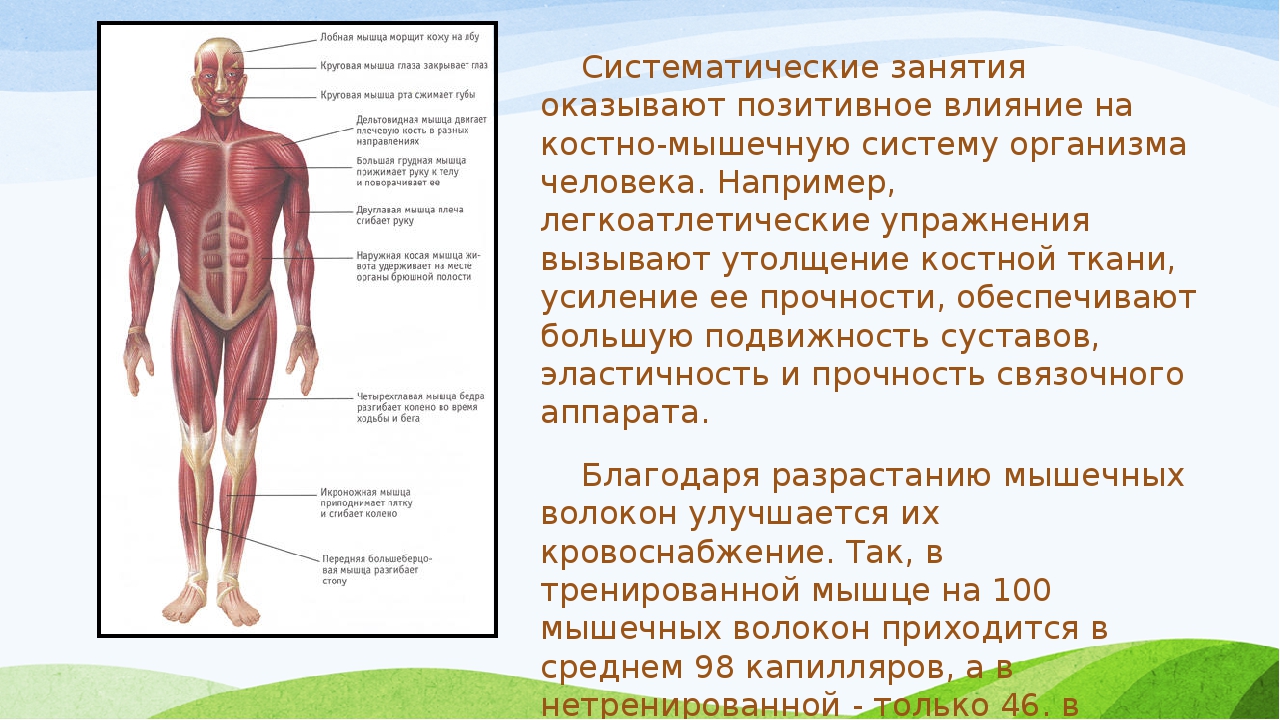 Если оценивать прикус в положении сидя, откинувшись назад, челюсть также смещается кзади. Такое положение отличается от обычного.
Если оценивать прикус в положении сидя, откинувшись назад, челюсть также смещается кзади. Такое положение отличается от обычного.
Фото №10,11 В эксперименте на крысах была изготовлена завышающая пломба на один боковой зуб с правой стороны. Через неделю peнтгенологическое обследование выявило деформацию всего позвоночника. После выравнивания окклюзии изготовлением пломбы на боковой зуб слева, форма позвоночника восстановилась.
Рисунок №12 Поскольку существует непосредственная связь между всеми частями скелетно-мышечной системы, несомненно, положение головы оказывает влияние на окклюзию, и может вызывать напряжение мышц шеи и спины.
Рисунок №13 При смещении нижней челюсти назад в результате аномалии прикуса в детстве или стираемости зубов со временем, шея перемещается вперед, и голова запрокидывается назад. Это, в свою очередь, значительно увеличивает нагрузку на мышцы шеи плечевого пояса. Поэтому у большинства людей с передним положением головы возникают боли в шейном отделе или напряжение мышц шеи.
Рисунок №14, 15 Часто смещение нижней челюсти при неправильном прикусе сопровождается ротацией первого и второго шейного позвонков, что ведет к выраженной деформации всего шейного отдела позвоночника.
Также, как изменения в положении челюсти могут вызывать изменения осанки и приводить к патологии в шейном отделе позвоночника, так и изменения в позвоночнике могут влиять на положение челюсти и окклюзию.
Роль дыхания и ночное апноэ
Рисунок №16 Смещение нижней челюсти назад часто приводит к сужению просвета дыхательных путей вследствие деформации шейного отдела позвоночника и смещения языка кзади, который следует за нижней челюстью. Для того, чтобы увеличить просвет дыхательных путей, мышцы еще больше перемещают шею вперед и запрокидывают голову назад.
При этом артерия, несущая кровь в головной мозг вдоль позвоночника, может резко перегибаться в области первого и второго шейных позвонков, вызывая нарушения кровоснабжения головного мозга.
Сужение дыхательных путей и нарушения кровоснабжения может вызывать состояние ночного апноэ – кратковременную остановку дыхания во время сна.
Повторяющиеся остановки дыхания приводят к значительному изменению метаболизма всего организма и, как следствие, могут стать одним из пусковых механизмов таких заболеваний как гипертония, сахарный диабет, инфаркт миокарда, инсульт и др. Поэтому нелеченное состояние ночного апноэ повинно в снижении продолжительности жизни на 20%.
Теперь Вы понимаете, что прикус гораздо сложнее, чем просто способ смыкания зубов! А неправильный прикус может быть причиной не только ранней потери зубов, но и вызывать такие серьезные патологии, как диабет и инфаркт миокарда и даже снижать продолжительность жизни.
Знакомство с мышечной системой
Мышечная система: факты и мифы
Когда вы думаете о мышцах, вы, вероятно, имеете в виду некоторых мускулистых людей, приятных для глаз. Знаете ли вы, что в вашем теле более 650 мышц? Звучит много, но у гусеницы даже больше, около 4000 мышц! Без мышц мы никогда не смогли бы двигаться. Основная функция мышечной системы — движение. Но знаете ли вы, что сокращения мышц производят 85% тепла нашего тела? Мышцы также защищают наши органы.Мышцы живота удерживают «кишки» и внутренние органы на месте. Некоторые мышцы имеют круглую форму, например та, которая окружает ваш рот для поцелуев или морщин. Другие круговые мышцы образуют сфинктеры, которые контролируют процесс дефекации или мочеиспускания. Одна важная мышца, известная как диафрагма, необходима для дыхания. Если ваша диафрагма перестанет работать, вы умрете в считанные минуты, потому что не сможете дышать.
Основная функция мышечной системы — движение. Но знаете ли вы, что сокращения мышц производят 85% тепла нашего тела? Мышцы также защищают наши органы.Мышцы живота удерживают «кишки» и внутренние органы на месте. Некоторые мышцы имеют круглую форму, например та, которая окружает ваш рот для поцелуев или морщин. Другие круговые мышцы образуют сфинктеры, которые контролируют процесс дефекации или мочеиспускания. Одна важная мышца, известная как диафрагма, необходима для дыхания. Если ваша диафрагма перестанет работать, вы умрете в считанные минуты, потому что не сможете дышать.
Давайте проверим ваши знания на некоторых общих фактах и мифах о мышечной системе.
Мышечная система и гомеостаз
Давайте посмотрим на мышечную систему в действии на следующем примере.
Сеймур — девятнадцатилетний пауэрлифтер и студент колледжа. Вы сидите рядом с Сеймуром на уроке психологии и познакомились с ним в этом семестре. В понедельник он садится рядом с вами и вздрагивает от боли, садясь на свое место. Вы спрашиваете его: «Что случилось?»
Вы спрашиваете его: «Что случилось?»
Он отвечает: «О, наверное, ничего, но у меня болит живот, особенно когда я поднимаю что-то тяжелое.”
«Разве вы не работаете в таверне« Темная лошадь »? И много тренироваться? Бьюсь об заклад, это должно быть жестко с тобой. Ты думаешь, ты потянул мышцу? » вы спросите его.
Сеймур отвечает: «Не знаю, я никогда раньше не чувствовал такой боли, мне было очень больно на работе, и я не мог так много поднимать в тренажерном зале».
«Почему бы нам не проверить это на моем iPhone, у нас еще есть 5 минут до начала занятий?» — отвечаете вы.
Сеймур предлагает зайти на WebMD.com, чтобы быстро взглянуть.«Хм, я не вижу ничего особенного в отношении боли в животе во время тренировки», — говорите вы ему. «Какие еще симптомы у вас имеются?»
«Ну, это немного неловко, но я заметил эту шишку в пупке, особенно во время подъема».
«У моего двоюродного брата раньше было нечто подобное, я думаю, это называлось грыжей, давай погуглим. О, блин, это похоже на то, что называется пупочной грыжей, там написано, что выпуклость может быть вашим кишечником! » ты шепчешь ему. «Сеймур, тебе лучше немедленно сходить к врачу, это никуда не годится.”
О, блин, это похоже на то, что называется пупочной грыжей, там написано, что выпуклость может быть вашим кишечником! » ты шепчешь ему. «Сеймур, тебе лучше немедленно сходить к врачу, это никуда не годится.”
Сеймур немного бледнеет и говорит: «Я позвоню маме сразу после урока».
Мышечная система в основном состоит из скелетных мышц, таких как те, которые покрывают переднюю часть живота и помогают сжимать брюшно-тазовую полость и органы пищеварения. Как мы видели на примере Сеймура, если эти мышцы ослаблены, они могут разделиться и позволить нижележащим органам выйти наружу. Постоянное воздействие на Сеймура высокого внутрибрюшного давления ослабило мышцы.Если кусок кишечника Сеймура выпячивается через мышцы живота, это может иметь серьезные последствия. Тонкий кишечник является важным компонентом пищеварительной системы, и если его ущипнуть, он может некротизироваться и погибнуть.
Есть два других типа мышечной ткани, которые необходимы для жизни: сердечная и гладкая мышца. Сердечная мышца находится в сердце, и ее ритмичное сокращение отвечает за ваше сердцебиение, а гладкие мышцы находятся во многих органах и кровеносных сосудах.Гладкая мускулатура является важной частью вашей сердечно-сосудистой системы (кровеносные сосуды), дыхательной системы (бронхиолы), пищеварительной системы (пищевод, желудок, тонкий и толстый кишечник), мочевыделительной системы (мочеточники и мочевой пузырь) и вашей репродуктивной системы (матка, семявыносящий проток).
Сердечная мышца находится в сердце, и ее ритмичное сокращение отвечает за ваше сердцебиение, а гладкие мышцы находятся во многих органах и кровеносных сосудах.Гладкая мускулатура является важной частью вашей сердечно-сосудистой системы (кровеносные сосуды), дыхательной системы (бронхиолы), пищеварительной системы (пищевод, желудок, тонкий и толстый кишечник), мочевыделительной системы (мочеточники и мочевой пузырь) и вашей репродуктивной системы (матка, семявыносящий проток).
Как указывалось ранее, скелетные мышцы выполняют множество важных функций в поддержании нашего гомеостаза. Сокращения скелетных мышц имеют решающее значение для выработки тепла для нашего тела. На самом деле, когда вам очень холодно, ваши скелетные мышцы увеличивают сокращение, чтобы генерировать больше тепла, чтобы согреть вас — это называется дрожью.Когда вы тренируетесь и ваши скелетные мышцы становятся более активными, вы производите больше тепла, и температура вашего тела повышается. Ваше тело потеет, чтобы охладиться.
Диафрагма — это скелетная мышца, которая сокращается, чтобы позволить нам вдыхать, мы выдыхаем, когда она расслабляется. Любое нарушение функции этой важной мышцы может быть фатальным. Диафрагма — важнейший орган дыхательной системы.
Ясно, что скелетные мышцы и мышечная ткань гораздо важнее, чем просто придание нам «баффового» тела!
Компоненты мышечной системы
Мышечная система состоит из скелетных мышц, соединенных с костями посредством сухожилий.Сухожилия — это тип плотной регулярной соединительной ткани, которая соединяет мышцы с костью. Мышцы живота также удерживаются широкими оболочками сухожилий, называемыми апоневрозами. В случае Сеймура мышца либо отделилась, либо оторвалась от апоневроза, что помогло скрепить мышечные волокна вместе.
Как уже упоминалось, существует три типа мышечной ткани. Скелетная мышца прикрепляется к костям, является произвольной и имеет полосатый (полосатый) вид. Сердечная мышца находится в сердце, непроизвольна и имеет поперечно-полосатую форму. Гладкая, иногда известная как висцеральная мышца, обнаруживается во многих органах и кровеносных сосудах, не имеет поперечной полосы, но возникает непроизвольно.
Гладкая, иногда известная как висцеральная мышца, обнаруживается во многих органах и кровеносных сосудах, не имеет поперечной полосы, но возникает непроизвольно.
Вся мышечная ткань состоит из мышечных клеток, известных как мышечные волокна. Эти волокна связаны и скрепляются соединительной тканью. Основная единица мышечной клетки — саркомер. Мышцы сокращаются, потому что саркомеры укорачиваются. Кальций необходим для правильного функционирования всех типов мышечной ткани. В этом модуле вы исследуете роль кальция в сокращении мышц.
Мышцы содержат много белков, в основном миозин и актин. Скелетная мышца находится под контролем нервной системы и не будет сокращаться, если нейронная команда не достигнет мышцы и не проинструктирует ее об этом. Нервная система взаимодействует со скелетными мышцами через химические посредники, известные как нейротрансмиттеры. Нейромедиатор, ацетилхолин, отвечает за стимуляцию скелетных мышц, поэтому они сокращаются. Процесс сокращения мышц происходит на клеточном уровне и будет изучен в этом блоке.
Мышечные волокна или клетки не все одинаковы. Есть клетки, которые специализируются на упражнениях на выносливость, они известны как медленно сокращающиеся волокна. Есть также клетки или волокна, которые лучше подходят для силового и спринтерского бега. Они утомляются намного быстрее и известны как быстро сокращающиеся волокна.
Когда вы изучаете мышечную систему, вам необходимо понимать микроскопическую анатомию и физиологию, чтобы вы могли узнать, как сокращаются мышцы. Вы также изучите анатомию мышц, их названия, расположение и функции в человеческом теле.Мышечная система — удивительная система. Наслаждайтесь путешествием по мере того, как вы его открываете!
МЫШЕЧНАЯ СИСТЕМА
МЫШЕЧНАЯ СИСТЕМА
МЫШЕЧНАЯ СИСТЕМА
Мышечная ткань тела составляет от одной трети до половины
массы тела среднего позвоночного.
Функции мышечной ткани:
движения и передвижения, благодаря его прямой связи с
система скелета
более тонких движений, связанных с поддержанием позы / вертикали
должность
помогает генерировать тепло из-за катаболических реакций, связанных с
с мышечной активностью (например, животные дрожат или увеличивают общее движение
при понижении температуры тела)
может быть преобразован в другие структуры, такие как электрические органы в
немного рыбы
В общую структуру мышечного волокна входят (рис. 10.2, п. 347) миофибриллы
10.2, п. 347) миофибриллы
(цепочки повторяющихся субъединиц), состоящие из двух видов филаментов:
тонких филаментов (состоящих из миозинов ) и толстых филаментов
(состоит из актина , тропомиозина и тропонина ), что
взаимодействуют путем связывания, создавая скользящее движение между нитями,
и это создает напряжение в мышечном волокне, ведущее к сокращению мышц.
Существует три общепризнанных типа мышечной ткани: гладкая, ,
сердечный
и скелет , каждый тип ткани с определенным расположением в теле,
клеточная организация (гистология) и общее действие мышечных волокон
(физиология)
Из-за множества функций мышц критерии классификации
мышцы включают:
1.Цвет
красный — сильно васкуляризован и богат миоглобином; сопротивляться усталости
белый — низкая васкуляризация и пониженное содержание миоглобина; быстрее
к усталости 2. Расположение
somatic — переместить кость или хрящ
висцеральный — контроль деятельности органов, сосудов или протоков 3.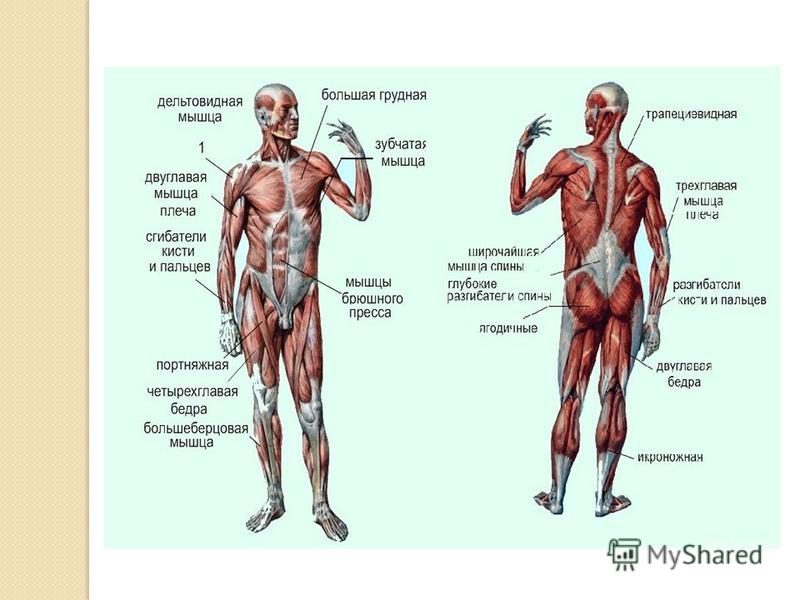 Контроль нервной системы
Контроль нервной системы
добровольно — под непосредственным сознательным контролем
принудительное — не 4.Эмбриональное происхождение
5. Общий вид под микроскопом
скелет — рис. 10.2, п. 347
сердечная — рис. 10.3, п. 348
гладкая — рис. 10.4, п. 348 Гладкая мышца
обнаружены выстилающие стенки кровеносных сосудов, внутренних органов (например,
пищеварительный тракт и матка), а также прикреплены к волоскам в
кожный покров.
два основных типа:
— унитарные гладкие мышцы самоинициализированные или миогенные
сокращение , чтобы помочь в поддержании ритмического движения органа
с чем это связано
— мультиединичная гладкая мышца имеет нейрогенное сокращение ,
который требует потенциалов действия, посылаемых нейронами, чтобы регулировать его действие. Сердечная мышца
находятся исключительно в мускулатуре сердечной стенки
в сердечной мышце, разветвление клеток увеличивает ее общее
связность и ячейки прочно объединены друг с другом через
интеркалированный
диски
Сердечная мышца не утомляется быстро, что является желательным признаком.
в мышцах, поддерживающих кровообращение
действие волокон сердечной мышцы демонстрирует смешанный контроль, так что
миогенный ритм сердца поддерживается нейрогенным контролем и
вся единица сердечной мышцы действует как синцитий или одиночный
функциональная единица Скелетная мышца
скелетные мышцы тесно связаны со скелетом и являются
используется в передвижении
каждое волокно скелетной мышцы также является синцитием из-за тесной связи
между сотовыми единицами
волокон тесно связаны с соединительными тканями и находятся под
произвольный контроль со стороны нервной системы.
| Гистология Полосатый? Форма Разветвленный? Расположение ядра Диски? Физиология | Гладкая Без бороздок Вынужденное | Сердечный полосатая Вынужденное | Скелетный полосатая Добровольное |
Общая терминология по мышцам
Как вы знаете из лабораторной работы, связано много уникальных терминов.
с мышечной системой, начиная от описания работы мышцы до
общая форма самой мышцы.
Термин «мышца» имеет как минимум два значения:
мышечная клетка или волокно — активный сократительный компонент:
мышечные клетки и их эндомизий
мышечный орган — весь орган: мышечные клетки плюс связанные
соединительные ткани, нервы, кровоснабжение Action
происходит за счет сокращения, которое создает напряжение в мышцах
так что он укорачивается и, таким образом, перемещает то, к чему прикреплен (будь то
кость, волосы или эпителий органа)
для скелетных мышц, каждая мышечная единица может быть описана на основе
ряд факторов, например, где находится основная мышца (живот , живот )
располагается, например, мышцы плеча, грудные мышцы, ягодичные мышцы,
и т.п.Мышца не прикрепляется непосредственно к кости сократительными мышечными волокнами.
— различные обертывания соединительной ткани выходят за концы
мышечные волокна для соединения с надкостницей кости:
сухожилие — шнуровидная насадка
апоневроз — плоский тонкий лист
фасция — тонкие плоские листы соединительной ткани, которые обертывают
и связать части тела вместе
raphe — соединение двух мышц на связке соединительной ткани
чтобы сформировать линию слияния, такую как linea alba
Основа для сокращения мышц:
мышца, не получающая нервного раздражителя, расслаблена или находится в
отдыхает
состояние — мягкая форма сохраняется за счет окружающих коллагеновых волокон
При воздействии нервных раздражителей за пределы порогового уровня мышц, сокращение
результатов и создается растягивающая сила , составляющая активную
государственный
прикрепленная кость и / или масса, которую необходимо переместить, представляет собой нагрузку
— действительно ли мышца сокращается, зависит от относительного баланса между
растягивающее усилие сжатия и перемещаемый груз (рис. 10.6, п.
10.6, п.
351)
Основные сократительные характеристики мышцы включают скорость ее достижения
максимальное напряжение и как долго он может выдерживать это напряжение
Напряжение и прочность напрямую связаны с количеством перемычек
между мышечными волокнами
в самом коротком положении, перекрытие нити мешает перемычке
образование и натяжение низкое (рис. 10.6а)
в самом длинном положении, нити очень мало перекрываются с небольшими перемычками
образования и слабое натяжение (рис.10.6b)
промежуточных отрезков обеспечивают максимальное поперечное перекрытие (рис. 10.6c)
Тонизирующие волокна:
относительно медленное сокращение и малое усилие
может выдерживать сокращение в течение длительных периодов времени
составляют большую часть осевого и аппендикулярного скелета Twitch (фазовые) волокна:
обычно производят быстрые сокращения, поэтому они часто составляют мышцы
используется для быстрого передвижения
медленное сокращение по сравнению с быстрым сокращением родственника, но медленное занимает примерно в 2 раза больше времени, чтобы
достичь максимальных сил Начало : конец мышцы, которая прикрепляется к более неподвижной части
скелета, который является проксимальным концом мышц конечностей
Место прикрепления : точка прикрепления мышцы, которая перемещает
чаще всего, когда мышца укорачивается и является самым дистальным концом мышц конечности
Для двуглавой мышцы живот лежит впереди плечевой кости, начало
клювовидный отросток лопатки, а место прикрепления — лучевой бугорок. Для трицепса источником являются задняя поверхность плечевой кости и
Для трицепса источником являются задняя поверхность плечевой кости и
инфрагленоидный бугорок лопатки, а место прикрепления — локтевый отросток
локтевой кости.
Действие скелетных мышц может быть:
антагонистический — противодействовать или сопротивляться действию другой мышцы
(например, бицепс и трицепс)
синергетический — работать вместе, чтобы произвести общий эффект (например,
как в действии сжатия кулака, в котором мышцы предплечья
и пальцы работают вместе)
Другие действия мышц включают:
Сгибатель — уменьшает угол в суставе
Extensor — увеличивает угол в суставе
Abductor — отводит кость от средней линии
Adductor — перемещает кость ближе к средней линии
Levator — производит движение вверх
Депрессор — производит движение вниз
Supinator — поворачивает ладонь вверх или кпереди
Пронатор — поворачивает ладонь вниз
Сфинктер — уменьшает размер отверстия
Tensor — делает часть тела более жесткой
Rotator — перемещает кость вокруг своей продольной оси.
Мы также можем описать мышцы на основе формы , например, в расположении
мышечных волокон:
Ремешковые мышцы — имеют параллельные волокна и широкие прикрепления.
(Терес майор)
Веретенообразные мышцы — параллельные волокна, но узкие сухожилия для прикрепления
(бицепс)
Перистые мышцы — расположенные по диагонали волокна, вставляющиеся сбоку
мышцы в сухожилие.(подлопаточная мышца)
Или Размер :
Максимус = самый большой
Minimus = самый маленький
Longus = самый длинный
Brevis = самый короткий Номер происхождения :
Бицепс = два начала
Трицепс = три начала
Квадрицепс = четыре начала Относительная форма
Дельтовидный = треугольный
Трапеция = трапеция
Serratus = зубчатый
Rhomboideus = ромбовидный или ромбовидный
Основные группы мышц соответствуют отделам, используемым для скелетных мышц.
система:
Осевые мышцы — мышцы туловища и хвоста рыб и четвероногих.
Бранхиометрические мышцы — также называемые висцеральными мышцами, например
как связанные с жабрами, челюстями и подъязычным аппаратом
Аппендикулярные мышцы — мышцы плавников рыб и мышцы конечностей
четвероногих
Мышцы возникают из трех эмбриональных источников:
мезенхима — рассредоточена по всему телу, образуя сглаживание
мышцы внутри стенок кровеносных сосудов и некоторых внутренних органов
висцеральный слой латеральной пластинки мезодермы — развивается в
гладкомышечные слои пищеварительного тракта и в стенки
сердце
параксиальная мезодерма или сомиты, и в частности слой миотома
сомита — основного источника скелетных мышц во время развития
— в области головы миотом не полностью сегментирован,
и вместо этого образует семь пар сомитомеров, которые будут производить мускулатуру
области головы (рис. 10.22а, стр. 365)
10.22а, стр. 365)
— остатки сомитов в теле развиваются в ствол и
аппендикулярные мышцы Гомологии
В процессе развития мускулов одни мышцы слились друг с другом, другие
разделились на отдельные новые мышцы, некоторые стали менее заметными,
а другие изменили точки привязанности и, следовательно, свою эволюцию
Гомологию мышц можно определить тремя способами:
— подобие вложений
— функциональное сходство
— нервная иннервация, обусловленная сохранением взаимоотношений между мышцами
и его нервное питание
Установление сходства может помочь сравнить разные группы мышц.
(краниальный, осевой и аппендикулярный) среди различных классов позвоночных
Черепные мышцы
Наружные мышцы глаза — шесть внешних мышц глаза, которые
прикрепляются к поверхности глаза и отвечают за перемещение глаза
в пределах орбиты (рис.10.23, стр. 366):
Дорсальный (верхний) косой Вентральный (нижний)
косой
Дорсальная (верхняя) прямая мышца брюшной стенки
(нижняя) прямая мышца
Медиальная прямая мышца
Боковая прямая мышца
Эти мышцы иннервируются глазодвигательным нервом.
У некоторых четвероногих также есть втягивающая луковица , которая притягивает глазное яблоко
дальше в орбиту, чтобы обеспечить покрытие мигательной мембраной
(отсутствует у людей)
Бранхиометрические мышцы — развиваются от миотомов каудально к
те, которые производят глазные мышцы
тесно связаны с висцеральным скелетом, поэтому они
используется как при дыхании, так и при кормлении.
выполняет функцию управления челюстью, открытия и закрытия
дыхальце (которое участвует в поступлении воды в жабры, когда рыба
поедание)
можно подразделить в зависимости от того, с какой висцеральной дугой они связаны
с:
| Жаберная дуга рыб Первая арка Вторая арка С третьей по седьмую арки | Мышцы Приводящая нижняя челюсть Межнижнечелюстная мышца Констриктор Леватор Констрикторы Леваторы | Действие закрывает челюсть сжимает горло сжимает жабры / глотку поднимает жаберные штанги сжимает жабры / глотку поднимает жаберные штанги |
cucullaris прикрепляется к последней жаберной дуге, но
связанный с грудным поясом
У четвероногих изменяется жаберная мускулатура одновременно с изменениями.
в висцеральном скелете, чтобы животные были более приспособлены к наземным
окружающая среда — привела к потере многих жаберных мышц
| Жаберная дуга тетрапода Первая арка Вторая арка | Мышцы Masseter Temporalis Крыловидные кости Дигастрический Мило подъязычная Платизма Грудинососцевидный Клейдомастоид | Действие закрывает челюсть закрывает челюсть Функция при движении губок открывает челюсть открывает челюсть перемещает кожу лица и шеи повернуть голову поворотная головка |
Наджаберные и гипобранхиальные мышцы — спинные и вентральные мышцы
связаны с областью головы и туловища, которые выполняют функции, связанные
с движением челюсти и языка
мышц рыб, связанных с кормлением и дыханием, включают:
— Coracoarcuals — открывает рот
— Коракомандибулярный — открывает рот
— Coracohyoid — помогает в кормлении
— Coracobranchial — помогает при глотании
мышцы четвероногих связаны с подъязычным аппаратом и
язык:
— Мышцы языка — hyoglossus, styloglossus, genioglossus
— Подъязычная мышца — краниально втягивает подъязычную кость
— Sternohyoid — втягивает подъязычную кость кзади
— Sternothyroid — втягивает гортань каудально
— эти мышцы также используются в речи и звуке у четвероногих.
Гомологии между жаберными и гипожаберными мышцами нескольких разных
Таксоны позвоночных представлены в Таблице 10.3.
Туловище / осевые мышцы
Осевая мускулатура туловища может функционировать либо
при движении или дыхании
Осевая мускулатура начинается с миотомов, разделенных миосептами, которые
затем делится на два региона:
эпаксиальные мышцы — мышцы тыльной части тела
гипаксиальные мышцы — мышцы вентральной части тела
которые разделены боковой перегородкой (рис.10.26, стр. 368) Рыбы
У рыб мышцы туловища остаются разделенными на складчатые мышечные сегменты.
или миомеры, которые делятся на миосепты
эти мышцы попеременно сокращаются, производя волнообразное движение.
который толкает рыбу по воде
внутренне эти мышцы остаются разделенными на спинных (эпаксиальных)
и вентральных (гипаксиальных) разрезов по боковой перегородке четвероногих
У четвероногих мышцы туловища больше работают при поддержании осанки,
движение головы и дыхание, а не движение, которое сместилось
аппендикулярным мышцам
Эпаксиальные мышцы скелета туловища четвероногих включают:
Longissimus dorsi — расширяет позвоночник.
Iliocostalis — сближает ребра
Multifidus spinae — расширяет позвоночный столб
Spinalis dorsi — расширяет позвоночник.
К гипаксиальным мышцам скелета туловища четвероногих относятся:
Пресс:
Rectus abdominis — сжимает живот
Внутренний косой — сжимает живот
Наружный косой — сужение живота
Внутренний косой — сужение живота
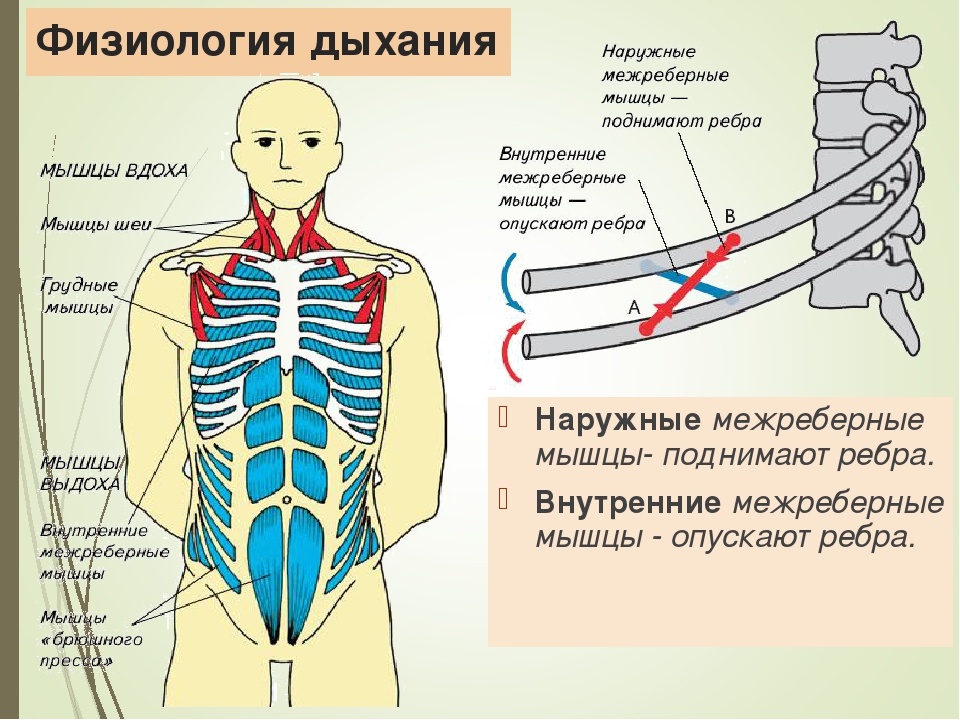
Дыхательные мышцы:
Serratus — ребра нарисуйте краниально)
Scalenus — сгибает шею)
Диафрагма — разделяет грудную / брюшную полость, функционирует в
дыхание
Межреберные мышцы — разгибание / втягивание ребер Аппендикулярные мышцы
Аппендикулярное развитие мышц происходит из сомитов в виде отростков
миотома сомита в зачаток конечности — миотомических зачатков в придатки
По мере роста зачатка конечности аппендикулярная мускулатура подразделяется на
мышечная масса, лежащая над аппендикулярным скелетом (спинные мышцы)
и масса, которая лежит ниже аппендикулярного скелета (вентральные мышцы)
Эти мышечные массы позже дифференцируются на несколько групп мышц.
в зависимости от типа организма
Рыбы
В общем, большая часть передвижения рыб зависит от действий
осевой мускулатуры, которая попеременно сокращается и расслабляется
производить волнообразные движения тела
Плавники (придатки) больше поддерживают стабильность,
торможение и маневрирование — таким образом, диапазон движения ласт намного больше
ограничен, чем у четвероногих конечностей
брюшные мышцы рыб идут в формирование отводящего
мышца , которая тянет плавники вентрально и краниально
спинные мышцы переходят в формирование приводящей мышцы
находится на постеродорсальной части плавника и перемещает плавник дорсально и
каудально Четвероногие
Аппендикулярная мускулатура четвероногих более сложна, чем у рыб
потому что конечности функционируют как в опоре, так и в движении
У четвероногих функция дорсальных и вентральных групп мышц.
— это , перевернутое на по сравнению с рыбами.
спинных мышц, которые у рыб отвечали за
приведение вместо этого отнимет или расширит придатки
вентральные мышцы, ранее использовавшиеся для отведения, вместо этого
используется для приведения или сгибания
| Грудная область Мышцы спины Мышцы груди Мышцы руки Тазовая область | Мышцы спины (разгибатели) Широчайшая мышца спины Максимум кожный Дельтоиды Мышцы спины Ягодичные мышцы | Вентральные мышцы (сгибатели) грудная мышца Бицепс Вентральные мышцы Приводящая мышца бедра Сгибатели пальцев |
Передвижение
Изучение передвижения завершает наше понимание скелетной
и мышечных систем, потому что он исследует функциональную взаимосвязь между
две системы, а также между организмом и окружающей его средой
Мы обсудим три важные категории передвижения: плавание,
земное передвижение и полет.
Плавание
Сначала мы можем различить плавающих позвоночных по тому, являются ли они первичными.
пловцы (виды, для которых плавание — единственный образец передвижения)
или вторичных пловцов (виды, которые полностью реадаптированы или
частично к водному образу жизни).
Некоторые общие требования к пловцам заключаются в том, что они должны:
1) уменьшить сопротивление воды движению движущегося
тело
2) продвигаются в относительно плотной среде
3) контроль вертикального положения в воде
4) сохранять ориентацию и управлять телом
Кроме того, пловцы-второстепенные спортсмены также должны пройти вторичную адаптацию.
их кровеносной, дыхательной и сенсорной системам, чтобы выдерживать высокие
давление и воздействие воды, которые влечет за собой плавание
Начальные пловцы — обычно волнообразные пловцы, использующие
мускулатура только плавников или плавники в сочетании с туловищем
и хвост, чтобы продвигаться по воде
Характеристики пловцов-первоклассников:
веретенообразное тело, жесткое удерживаемое за счет сильного сочленения
позвоночник
сегментированных миомеров, которые позволяют отдельным мышечным единицам проявлять силы
по всей стороне тела
Покровный покров, прочно прикрепленный к подлежащей мускулатуре посредством
соединительная ткань для увеличения компактности тела
Мускулатура, скелетная система и покровы составляют единое целое.
что помогает оптимизировать животное и уменьшить сопротивление при движении
вода
Второстепенные пловцы — обычно колеблющиеся пловцы, которые двигаются
сами по воде с лопастными движениями придатков
придатков также можно преобразовать в лямки или ласты для помощи
в движении
обычно имеют хорошо развитую аппендикулярную мускулатуру
может уменьшить сопротивление давления вокруг тела за счет временной оптимизации
сами при движении по воде Наземное движение
Что касается наземного передвижения, существует множество различных режимов, которые
животное может использовать:
Cursorial — четвероногие, которые путешествуют далеко или быстро по суше.Бегущие животные обладают относительно удлиненным телом, в котором позвоночные
Колонна увеличивает шаг бега, растягиваясь для увеличения шага вперед
движущая сила
Saltatorial — четвероногие, которые прыгают или прыгают. Сальтаторские животные
имеют тела, в которых вес перенесен на задние лапы, лапы
мощный и прочный, а центр масс выровнен с
крестец
Scansorial — четвероногие, умеющие лазать.Сканирующие животные
укрепили грудную мускулатуру и придатки, а также изменили фаланги
для цепляния за вертикальные поверхности
Fossorial — четвероногие, которые умеют копать и живут в некоторой степени
подземное существование. Ископаемые животные обладают очень гибкими позвоночными
колонны, сильная грудная мускулатура и модифицированные фаланги для копания
Каждый из этих различных образов жизни требует кардинальных изменений в
скелетная система (с точки зрения частей тела, которые получают
наибольшее напряжение) и связанная с ним мускулатура
Животные, передвигающиеся на двух ногах или сканирующие, имеют позу ступней
более стопоходящий, при котором подошвы ступней расположены ровно
на земле.Напротив, у большинства бегающих животных более пальцевидная мышца.
поза, в которой запястье и лодыжка отрываются от земли, а
животное ходит на пальцах. Или животное может быть очень длинноногим,
и ходите только по кончикам цифр так, чтобы конечный конец
цифра видоизменяется, образуя копыто, а другие цифры теряются, осанка
называется унгулиграда.
Рейс
Тетраподы, которые летают, могут летать тремя разными способами.
зона для предотвращения случайного падения
площадь и пройти большее горизонтальное расстояние по воздуху
воздух
Четвероногие животные, которые используют активный полет, например птицы, имеют грудные придатки.
которые уменьшены до однозначного числа и имеют высокоразвитую грудную мускулатуру
необходимо для поддержания активного полета
У птиц первичные летные мышцы берут начало на вентральной части .
поверхность расширенной килевой грудины:
pectoralis — депрессор крыла
supracoracoideus — поднимающий крыло
У летучих мышей летные мышцы связаны с движением
плечевая кость и лопатка и расположены на стороне грудной клетки:
основные депрессоры крыла:
— грудная мышца
— подлопаточная мышца
— передняя зубчатая мышца
леваторы основного крыла:
— дельтовидная мышца
— трапеция
— спинатус
Определения
Отводящий — отводит кость от средней линии
Adductor — перемещает кость ближе к средней линии
Антагонистическое — состояние, при котором мышца сопротивляется или сопротивляется действию.
другой мышцы
Апоневроз — листовое сухожилие мышцы
Cursorial — четвероногие, которые путешествуют далеко или быстро по суше.
Депрессор — производит движение вниз
Digitigrade — поза, в которой запястье и лодыжка отведены от
земля и животное ходит на пальцах
Эпаксиальный — относится к структурам, лежащим выше или рядом с позвонком.
ось
Extensor — увеличивает угол в суставе
Фасции (фасции) — листы соединительной ткани, которые лежат под кожей или покрывают оболочку.
группы мышц
Flexor — уменьшает угол в суставе
Полет — использование крыльев для активного поддержания движения в воздухе
Ископаемые — четвероногие, умеющие копать, и немного живущие.
подземное существование
Скольжение — использование широких перепонок, прикрепленных к конечностям для увеличения поверхности
области и пройти большее горизонтальное расстояние по воздуху
Гипаксиальный — относится к структурам, лежащим вентрально по отношению к позвоночному.
ось
Insertion — точка прикрепления мышцы, которая больше всего движется
когда мышца укорачивается, или самый дистальный конец мышц конечностей
Levator — производит движение вверх
Начало — конец мышцы, которая прикрепляется к более фиксированной части
скелет, который является проксимальным концом мышц конечностей
Колеблющиеся пловцы — продвигаются по воде с помощью весла.
движения придатков
Парашютный спорт — использование конечностей и тела для увеличения общей площади поверхности
прервать случайное падение
Плантиградская поза — поза, в которой подошвы ступней расположены ровно.
на земле во время передвижения
Пловцы-основатели — виды, для которых плавание является единственным
движение
Пронатор — поворачивает ладонь вниз
Raphe — соединение двух мышц в связке соединительной ткани с образованием
линия слияния, такая как linea alba
Вращатель — перемещает кость вокруг продольной оси
Saltatorial — четвероногие, которые прыгают или прыгают
Scansorial — четвероногие, умеющие лазать
Пловцы среднего уровня — виды, полностью или частично реадаптировавшие
к водному образу жизни от наземной жизни
Сфинктер — уменьшает размер отверстия
Supinator — поворачивает ладонь вверх или кпереди
Синергетический — состояние, при котором мышцы работают вместе, чтобы производить
общий эффект
Tensor — делает часть тела более жесткой
Пловцы на волнах — используйте только мускулатуру ласт или ласты.
в сочетании с туловищем и хвостом, чтобы продвигаться через
вода
Унгулиграда — локомоторная поза, используемая длинноногими четвероногими, которые
ходите только по кончикам цифр таким образом, чтобы конечный конец цифры
изменяется, образуя копыто, и другие пальцы теряются
Мышечная система: действия и физиологические процессы
Мышцы состоят из мышечных волокон
Мышечная система состоит из специализированных мышечных клеток или мышечных волокон, которые называются миофибрами . Myo- происходит от греческого слова mys , что означает «мышь» или «мышца». (По-видимому, греки думали, что мышцы напоминают мышей, движущихся под кожей.)
Каждое мышечное волокно наполнено белками, которые вместе создают сокращения. Мышечные сокращения — это индивидуальные клеточные действия, которые вызывают движение в нашем теле. Эти стеки сократительных белков заставляют определенные типы мышц выглядеть полосатыми.
Миофибры организованы в пучки, которые образуют мышцы , основные органы мышечной системы.
Функции мышечной системы
Мышцы прикреплены к костям, органам и кровеносным сосудам и отвечают за жизненно важные задачи движения, поддержание осанки и равновесия, а также создание тепла для тела.
Создание движения
Произвольное движение — это движение, которое вы осознанно осознаете, например, касание пальцами ног и прыжки вверх и вниз. Это происходит, когда мышцы тянут кости. Мышцы соединены с костями посредством сухожилий . Когда мышцы сокращаются, сухожилия тянут кости, а суставы действуют как рычаги, перемещающие ваше тело во всех направлениях.
Мышцы работают парами. Одна группа мышц, например бицепс, будет сокращаться, чтобы кости двигались в одном направлении.Затем эта группа мышц расслабится, в то время как ее партнерская группа, в нашем примере трицепс, сокращается, чтобы вернуть ваши кости в противоположном направлении.
Непроизвольное движение — это движение, о котором вы не подозреваете, например, пищеварение и дыхание. Это вызвано мышцами, выстилающими ваши органы.Например, во время глотания мышцы пищевода последовательно сокращаются и расслабляются, выталкивая пищу вниз по направлению к желудку. Как только пища попадает в желудок, мышцы стенки желудка сокращаются и расслабляются, смешивая пищу с пищеварительными химическими веществами.
Поддержание осанки и равновесия
Ваши постуральные мышцы , которые представляют собой мышцы живота, спины и бедер, устанавливают и поддерживают правильную вертикальную осанку. Эти мышцы действуют как корсет, удерживая ваши органы и верхнюю часть тела на месте.Поддержание формы и сильных постуральных мышц важнее для вашего здоровья в целом, чем вы, вероятно, думаете. Он уменьшает боль в спине, поддерживает ваши легкие и помогает дышать, а также позволяет вашему телу становиться выше и прямее.
Кроме того, все мышцы постоянно обеспечивают обратную связь с мозгом относительно положения вашего тела и частей тела. Мышцы, в свою очередь, получают сигналы, чтобы постоянно вносить незначительные изменения, позволяя вам сохранять равновесие и осанку.Фактически, ваши мышцы никогда не расслабляются полностью. Это называется мышечным тонусом, он предотвращает травмы наших мышц и суставов и позволяет нам оставаться в стабильном положении, даже когда мы спим.
Производство тепла
Нашему телу необходимо поддерживать постоянную температуру, чтобы нормально функционировать. Большая часть этого тепла, около 70-80%, производится нашими мышцами. Когда клетки мышц сокращаются, они выделяют тепло. Вот почему во время тренировки вам становится жарко и потеет.Ваше тело использует это, чтобы регулировать температуру. Когда вы начинаете становиться слишком холодными, ваш мозг посылает сигнал мышцам, чтобы они быстро сокращались и расслаблялись, выделяя тепло. Это известно как дрожь .
Вы также могли заметить, что чувствуете усталость и вялость в очень жаркие дни. Это потому, что ваш мозг сигнализирует вашему телу замедлиться, расслабиться и перестать сокращать эти мышцы. Меньшее сокращение мышц означает меньшее тепловыделение, что важно, когда вам становится слишком жарко.
Типы мышц
Существует три типа мышечной ткани: скелетная или поперечно-полосатая мышца, гладкая мышца и сердечная мышца.
Скелетная мышца прикрепляется к костям и отвечает за движение вашего скелета. Этот тип мышц иногда называют поперечно-полосатыми мышцами из-за его полосатого вида. «Полосатый» — это просто причудливое слово для обозначения «полос» или «бороздок».Эти мышцы находятся под произвольным или сознательным контролем.
Гладкая мышца находится в стенках полых органов вашего тела, таких как кровеносные сосуды, кишечник и мочевой пузырь. Они называются «гладкими», поскольку в них отсутствуют бороздки или бороздки, которые можно найти на тканях скелетных и сердечных мышц. Эти мышцы непроизвольно сокращаются и расслабляются, что означает, что они не находятся под сознательным контролем. Гладкие мышцы сокращаются медленными ритмичными движениями и обладают высокой эластичностью.Например, мочевой пузырь человека при необходимости может увеличиваться более чем в три раза по сравнению с исходным размером. Матка превосходит это, увеличиваясь с апельсина до размера арбуза к концу беременности.
Сердечная мышца , обнаруженная в стенках сердца, непроизвольно сокращается, как гладкая мышечная ткань, но имеет поперечно-полосатую форму, как ткань скелетных мышц. (Между прочим, «сердечный» происходит от греческого слова kardia , что означает «сердце».) Сокращения сердечных мышц ритмичные и сильные, заставляя кровь двигаться по всему телу.
Группы мышц
Основные группы мышц можно разделить на категории по месту их расположения на вашем теле. Мышцы головы и шеи отвечают за мимику, движение глаз, жевание, глотание и стабилизацию головы. Мышцы туловища, такие как грудь, живот и спина, перемещают позвонки, поддерживают осанку и участвуют в дыхании. Верхние конечности — это мышцы рук, которые приводят в движение плечи, предплечья, предплечья, запястья и кисти рук.Нижние конечности — это мышцы ваших ног, они двигают ноги, лодыжки и ступни.
Краткое содержание урока
Мышечная система — это система органов, состоящая из всех мышц тела. Он отвечает за произвольные и непроизвольные движения, поддерживает осанку и равновесие, а также выделяет тепло. Мышечная система состоит из специализированных мышечных клеток, называемых миофибрами .Миофибры содержат белки, которые работают вместе, создавая сокращения, клеточные действия, которые вызывают движение в нашем теле. Миофибры собраны в пучки, образуя мышц и основных органов мышечной системы.
Есть три типа мышечной ткани: скелетная мышца, гладкая мышца и сердечная мышца. Скелетная мышца прикреплена к костям и отвечает за сознательные, произвольные движения вашего скелета. Гладкая мышца находится в стенках полых органов вашего тела, таких как кровеносные сосуды, желудочно-кишечный тракт и диафрагма, и отвечает за бессознательные непроизвольные движения. Сердечная мышца , обнаруженная в стенках сердца, непроизвольно сокращается, как гладкая мышечная ткань, но имеет поперечно-полосатую форму, как ткань скелетных мышц. Сильные ритмичные сокращения сердечной мышцы отвечают за перекачку крови по всему телу.
Мышечная система — определение, функции и составные части
Определение
Мышечная система — это совокупность тканей тела, обладающих способностью изменять форму. Мышечные клетки соединяются вместе и в конечном итоге с элементами скелетной системы.Когда мышечные клетки сокращаются, создается сила, когда мышцы тянутся к скелету.
Обзор
Актин и миозин — основные белки, используемые в мышечных клетках для сокращения. На изображении ниже актин показан зеленым цветом, а миозин — фиолетовым. Эти два компонента используют АТФ, чтобы притягивать друг друга. Они прикрепляются к каждой стороне ячейки, что укорачивает ячейку, когда они проходят друг мимо друга.
Как видно на рисунке ниже, мышечная система сокращается, когда энергия АТФ применяется к миозиновым головкам миозинового белкового волокна.Голова выпускает актин, тянется вперед и снова захватывает актин. Это перемещает белковые нити и сокращает волокна. В зависимости от мышечной клетки могут использоваться разные формы актина и миозина. В некоторых организмах используются совершенно разные белки.
Сокращение скелетных мышц
Мышечная система основана на скоординированном действии миллионов актиновых и миозиновых нитей, тянущихся в одном направлении в одно и то же время. Для достижения этой координации мышцы иннервируются нервной системой. Нервные сигналы, исходящие из мозга, направляются к определенным мышцам, позволяя организмам стимулировать определенные мышечные ткани для выполнения скоординированных действий, таких как бег, плавание и полет.
Функция мышечной системы
Механизм
Самая очевидная функция мышечной системы — движение. Организмы применили множество методов, чтобы использовать сократительную функцию мышечной системы для передвижения в окружающей среде. Самые основные движения рыбы включают последовательное сокращение мышц на противоположных сторонах тела.Это действие продвигает их по воде.
У организмов с конечностями, сухожилия и другие соединительные ткани используются для прикрепления мышц к суставам и скелету. Скелеты могут быть внутренними, как человеческие скелеты, или они могут быть внешними, как экзоскелет крабов. Нервная система координирует сокращение мышечной системы, чтобы синхронизировать движения конечностей. Такие животные, как гепард, рыба-меч и летучая мышь, развивают скорость выше 60 миль в час или более только благодаря силе своих мускулов.
Тираж
Вторая и менее очевидная функция мышечной системы — способствовать кровообращению. Ткани висцеральных и сердечных мышц окружают кровеносные и лимфатические сосуды, которые несут важные питательные вещества и кислород к клеткам тела. Сердечная мышца составляет сердце и обеспечивает основную силу для крови, перемещающейся по телу.
Крупные артерии и вены связаны с мышцами, которые могут сокращаться или расслабляться, чтобы контролировать кровяное давление.Действия крупных скелетных мышц также помогают перекачивать кровь и лимфатическую жидкость по всему телу. Когда вы тренируетесь и сокращаете большие и маленькие мышцы, они отталкивают сосуды в сторону, что работает как насос, перемещая жидкости по вашему телу.
Пищеварение
Подобно своей способности перемещать жидкости по сосудам в системе кровообращения, мышечная система также помогает перемещать пищу через пищеварительную систему. Большинство органов пищеварения окружены гладкой мышечной тканью. Хотя ткань не может быть сокращена произвольно, как скелетные мышцы, она контролируется подсознательно. Когда пища должна перемещаться по кишечнику, мышцы сокращаются синхронно, волнообразно через пищеварительную систему. Эти волнообразные мышечные сокращения называются перистальтикой .
частей мышечной системы
В отличие от других систем органов, мышечная система делится на разные типы тканей, которые входят в состав различных органов тела.
Схема мышечной системы
Скелетные мышцы
Поперечно-полосатая мышца, или Скелетная мышца, — ткань, наиболее часто связанная с мышечной системой.Этот тип мышц прикрепляется к скелету и перемещает конечности и тело организма. Системы скелетных мышц состоят из поперечно-полосатых мышц , которые имеют отдельные группы белков в каждой миофибрилле . Когда этим белкам дается энергия, они скользят друг мимо друга, стягивая концы каждой мышечной клетки вместе. Саркомеры , или функциональные единицы актина и миозина, образуют полосатость, которую можно увидеть в поперечно-полосатых мышцах. Это можно увидеть на изображении ниже.
Скелетная мышца
Висцеральная мышца
Напротив, клетки висцеральных мышц не содержат этих резких полос белка, а волокна актина и миозина работают по-разному. Вместо толстых волокон, проходящих через клетку, висцеральная мышца окружена сеткой из актиновых и миозиновых волокон, которые сжимают клетку при сокращении. Это можно увидеть на изображении ниже. По этой причине висцеральная мышца также известна как гладкая мышца .
Сокращение гладких мышц
Сердечная мышца
Сердечная мышца, , которая окружает камеры сердца, полосатая, как скелетная мышца, но клетки соединены с соседними клетками, что создает большее сократительное движение для перекачивания крови.
Структура мышечной системы
В целом, мышечная система имеет базовую структуру, которая позволяет мышцам двигать конечностями и создавать силу. Мышца всегда расположена между двумя костями и связана с костями посредством сухожилий, которые представляют собой волокнистые и гибкие ткани, которые могут прикрепляться к костям. Действие укорачивания каждой отдельной клетки заставляет мышцу сокращаться в целом. Это натягивает сухожилия с каждой стороны мышцы, создавая нагрузку на кости. Кости, если они соединены в сустав, могут двигаться в ответ на эту силу.
Некоторые кости неподвижны, что позволяет мышцам натягивать их. Так обстоит дело с такими мышцами, как диафрагма, которая связана с двумя неподвижными костями. Когда диафрагма сжимается, она вытесняет воздух из грудной полости, потому что вся сила направлена вверх.
Вместе, множество различных типов и соединений мышц функционируют, чтобы дать вашему телу полный диапазон движений. Таким образом, многие мышцы пересекаются друг с другом или находятся на противоположных сторонах кости, чтобы перемещать ее в разных направлениях.
Соединения мышечной системы с нервной системой
Скелетная мышца в основном связана с соматической нервной системой , которая контролируется произвольными импульсами мозга. С другой стороны, сердечные и висцеральные мышцы контролируются в основном вегетативной нервной системой , которая контролирует подсознательные действия организма. Разделение этих нервных систем гарантирует, что вегетативные функции, такие как дыхание и пищеварение, продолжаются, когда животное движется и ищет больше пищи.
Различия в тканях мышечной системы связаны с их очень разным использованием. Скелетные мышцы должны быстро выполнять большой объем работы, поэтому они состоят из поперечно-полосатых мышечных клеток, которые могут произвольно сокращаться. Гладкая мышечная ткань висцеральных тканей имеет меньше митохондрий, производящих энергию. Эти ткани просто используются для сокращения полых органов и перемещения жидкости внутрь. Желудок, кишечник и кровеносные сосуды выстланы висцеральными мышцами. Сердечная мышца имеет поперечно-полосатую форму, потому что ей необходимо создавать большую силу, хотя она не контролируется добровольно.
Заболевания мышечной системы
Заболевания мышечной системы подразделяются на множество категорий. Могут произойти простые травмы мышечной системы, например, разрыв мышцы или растяжение лодыжки. Такие заболевания, как тендинит, могут возникнуть при неоднократном растяжении сухожилия. Однако есть несколько мышечных заболеваний, не вызванных напряжением или повреждением реальных мышечных клеток.
Мышечная дистрофия — это генетическое заболевание, поражающее мышечную систему. Начиная с 2-6 лет заболевание приводит к стойкому ослаблению мышц по всей анатомии.Это продолжается до конца жизни больного человека. Как правило, люди с мышечной дистрофией доживают только до подросткового возраста или до двадцати пяти лет.
Волчанка — еще одно заболевание, поражающее мышечную систему. Симптомы волчанки включают сыпь в форме бабочки на лице, а также отек и воспаление кожи, мышц и суставов. Волчанка — это аутоиммунное заболевание, то есть причина заболевания — иммунные клетки в вашем теле, вырабатывающие антитела против собственных белков вашего тела.
Викторина
ФУНКЦИИ И СВОЙСТВА МЫШЕЧНОЙ ТКАНИ
ФУНКЦИИ И СВОЙСТВА МЫШЕЧНОЙ ТКАНИ
Функции мышечной ткани
- Движение: Скелет нашего тела придает достаточно жесткости нашему телу, чтобы скелетных мышц могли тянуть и дергать его, что приводит к движениям тела, таким как ходьба, жевание, бег, подъем, манипулирование объектами руками , и ковырять в носах.
- Поддержание осанки: Без особого сознательного контроля наши мышцы генерируют постоянную сократительную силу, которая позволяет нам сохранять прямое или сидячее положение, или позу .
- Дыхание: Наша мышечная система автоматически управляет движением воздуха внутрь и наружу.
- Выработка тепла: При сокращении мышечной ткани выделяется тепло, которое необходимо для поддержания температурного гомеостаза.Например, если температура нашего тела падает, мы дрожим, чтобы генерировать больше тепла.
- Связь: Мышечная ткань позволяет нам говорить, жестикулировать, писать и передавать свое эмоциональное состояние, делая такие вещи, как улыбка или хмурый взгляд.
- Сужение органов и кровеносных сосудов: Питательные вещества проходят через наш пищеварительный тракт, моча выводится из организма, а секреты выводятся из желез за счет сокращения гладких мышц . Сужение или расслабление кровеносных сосудов регулирует кровяное давление и распределение крови по телу.
- Перекачивание крови: Кровь движется по кровеносным сосудам, потому что наше сердце без устали получает кровь и доставляет ее ко всем тканям и органам тела .
- Это не полный список. Среди множества возможных примеров можно отметить тот факт, что мышцы помогают защитить хрупкие внутренние органы, заключая их, а также имеют решающее значение для поддержания целостности полостей тела.Например, у плода с не полностью сформированной диафрагмой брюшное содержимое грыжи (выступает) вверх в грудную полость, что препятствует нормальному росту и развитию легких. Несмотря на то, что это неполный список, понимание некоторых из этих основных мышечных функций поможет вам в дальнейшем.
Свойства мышечной ткани
Все мышечные клетки имеют несколько общих свойств: сократимость, возбудимость, растяжимость и эластичность:
- Сократимость — это способность мышечных клеток сильно сокращаться.Например, чтобы согнуть (уменьшить угол сустава) локоть, вам нужно сократить (укоротить) двуглавую мышцу плеча и другие мышцы-сгибатели локтя в передней части руки. Обратите внимание, что для того, чтобы вытянуть локтя на , мышцы-разгибатели задней части руки должны сократиться. Таким образом, мышц могут только тянуть, но не толкаться.
- Возбудимость — это способность реагировать на стимул, который может быть доставлен двигательным нейроном или гормоном.
- Растяжимость — это способность мышцы растягиваться. Например, давайте пересмотрим наше движение сгибания локтя, которое мы обсуждали ранее. Чтобы иметь возможность сгибать локоть, мышцы-разгибатели локтя должны разгибаться, чтобы можно было сгибать. Отсутствие растяжимости известно как спастичность .
- Эластичность — это способность отскакивать или возвращаться к исходной длине мышцы после растяжения.
** Вы можете использовать кнопки ниже, чтобы перейти к следующему или предыдущему чтению в этом модуле **
Распечатать эту страницу
границ | Внеклеточный матрикс скелетных мышц — что мы знаем о его составе, регуляции и физиологической роли? Повествовательный обзор
Введение
Скелетные мышцы являются важным компонентом состава тела человека, обычно составляя более 40% и 30% общей массы тела у мужчин и женщин, соответственно (Kim et al., 2002). Наиболее очевидная функция скелетных мышц — генерировать силы, необходимые для поддержания вертикальной позы и движения. Однако скелетные мышцы также играют важную роль во многих других физиологических процессах, включая термогенез (Rowland et al., 2015), метаболизм (Baskin et al., 2015) и секрецию многочисленных пептидов для связи с другими тканями (Pedersen and Febbraio , 2012). Таким образом, укрепление и поддержание здоровья скелетных мышц имеет жизненно важное значение.Хотя в последние годы фармакологические имитаторы упражнений вызывают все больший научный интерес (Fan and Evans, 2017), именно физические упражнения по-прежнему считаются наиболее мощным и универсально применимым инструментом для этих целей.
За последние десятилетия были выполнены тысячи тренировочных исследований в попытке определить методы упражнений, наиболее подходящие для увеличения размера мышц и улучшения их функциональных характеристик в разных когортах (например, на момент написания этой рукописи Pubmed дал больше более 24 000 результатов по операторам поиска «упражнения» и «сила мышц»).Результаты этих исследований послужили источником вдохновения для различных руководств по назначению упражнений, вероятно, наиболее известными из которых являются стенды с позициями, публикуемые и обновляемые через нерегулярные промежутки времени Американским колледжем спортивной медицины (2009), Garber et al. (2011). Большинство исследований основывают свою оценку эффективности тренировочных вмешательств на исследовании сократительных мышечных клеток. Часто изучаемые параметры включают размер мышц, измеренный на уровне органа (Fisher et al., 2011) или на клеточном уровне (Schoenfeld, 2010), распределение типов волокон (Adams et al., 1993), архитектуру (Aagaard et al., 2001), а также нервный импульс к мышцам (Folland and Williams, 2007).
Несмотря на обилие информации о пластичности скелетных мышц, часто упускается из виду тот факт, что мышечные волокна встроены во внеклеточный матрикс (ЕСМ), состоящий из сетки коллагеновых компонентов, а также смеси других макромолекул, таких как различные гликопротеины и протеогликаны. Недавние исследования показали, что ECM играет важную роль в развитии (Thorsteinsdóttir et al., 2011), рост (Fry et al., 2017) и восстановление мышц (Calve et al., 2010), а также передача сократительной силы (Street, 1983). Несмотря на то, что доказательства, демонстрирующие пластичность ЕСМ, существуют, лишь в небольшом количестве исследований сообщается о его реакциях на различные формы тренировок, предполагая, что физиологическая роль ЕСМ еще не полностью оценена специалистами по физическим упражнениям. Стремясь стимулировать дальнейшие исследования тренировочных реакций неконтрактильных компонентов скелетных мышц, мы предлагаем обзор текущего состояния знаний о составе, структуре и регуляции внеклеточного матрикса, его физиологической роли, нарушениях регуляции, связанных со старением и метаболизмом. расстройства, а также адаптации к физическим нагрузкам.
Состав и структура ECM скелетных мышц
ECM скелетных мышц представляет собой сложную сеть, состоящую из коллагенов, гликопротеинов, протеогликанов и эластина (Takala and Virtanen, 2000; Halper and Kjaer, 2014). Коллагены образуют сеть внутримышечной соединительной ткани (IMCT), то есть центральные фиброзные компоненты ECM. IMCT обычно изображается организованной в виде трех слоев: (i) эндомизий, представляющий самый внутренний слой, который охватывает отдельные мышечные волокна, (ii) перимизий, связывающий группы мышечных волокон, и (iii) эпимизий, охватывающий всю мышцу.Большая структурная сложность сети ММКИ, подтвержденная сканирующими электронными микрофотографиями, предполагает, что эта традиционная классификация может быть упрощенной и что организация ECM мышц более высокого порядка все же нуждается в определении (Gillies and Lieber, 2011). Исследования фасциальных тканей также рассматривают слои IMCT как часть сложной системы взаимосвязанных и переплетенных соединительных тканей, которая «окружает, переплетается между собой и проникает во все органы, мышцы, кости и нервные волокна, обеспечивая тело функциональной структурой, и обеспечение среды, которая позволяет всем системам организма работать интегрированно » (Adstrum et al., 2017; Stecco et al., 2018). Эта система, которую обычно называют фасциальной системой, все чаще признается важной мишенью в спортивной медицине (Zügel et al., 2018).
IMCT содержит различные формы коллагенов, наиболее распространенными из которых являются типы I и III (Duance et al., 1977; Light and Champion, 1984; Gillies and Lieber, 2011; McKee et al., 2019). Эндомизий взаимодействует с сарколеммой миофибр на специальной базальной мембране, которая состоит в основном из коллагена IV типа и ламинина (Sanes, 1982; Martin and Timpl, 1987; Kjaer, 2004).Было обнаружено, что концентрация этих двух компонентов различается в зависимости от типа мышечных волокон, при этом медленно сокращающиеся волокна характеризуются значительно более высокими концентрациями коллагена IV, но более низкими концентрациями ламинина (Kovanen et al., 1988). Ламинин, в свою очередь, служит лигандом для двух сарколеммальных рецепторов — гликопротеинового комплекса, ассоциированного с дистрофином и интегрина α7β1 (Grounds et al., 2005), — расположенных на костамерах, которые представляют собой мембранные белковые структуры, выровненные в соответствии с Z- диски миофибрилл.Считается, что интегрины действуют двунаправленным образом, позволяя внутриклеточным сигнальным молекулам регулировать внешнюю адгезию (передача сигналов «Inside-Out») и передавать внешние стимулы для воздействия на клеточные процессы (передача сигналов «Outside-In») (Boppart and Mahmassani, 2019) . Таким образом, явный стресс цитоскелета индуцирует внутриклеточное связывание белков, таких как талин, винкулин или киндлин, что приводит к изменению конформации рецептора интегрина и позволяет внеклеточным доменам рецептора распространяться в направлении белков внутри ЕСМ.Кроме того, лиганды интегринов из внеклеточного пространства, такие как ламинин, коллаген или фибронектин, способствуют формированию высокоаффинного вертикального состояния, что приводит к усилению связывания с белками ЕСМ и кластеризации интегринов, особенно вдоль комплексов фокальной адгезии (Boppart and Mahmassani, 2019) . Гликопротеиновый комплекс, связанный с дистрофином, является еще одним важным фактором в обеспечении механической связи между сократительными компонентами скелетных мышц (т.е.актином) и взаимосвязанными слоями IMCT (Ervasti, 1993; Peter et al., 2011). Основные компоненты, связывающие сократительные элементы мышцы с интерстициальным матриксом, показаны на рисунке 1.
Рисунок 1. Основные компоненты внеклеточного матрикса скелетных мышц и его связь с сократительными компонентами мышцы.
Суперсемейство коллагена состоит в общей сложности из 28 различных членов, из которых типы I, III, IV, V, VI, XII, XIII, XIV, XV, XVIII и XXII, как было показано, присутствуют в зрелых скелетных мышцах в гене и / или уровень белка (Таблица 1).Формирующие фибриллы типы I и III, безусловно, наиболее многочисленны, и протеомные исследования показывают, что они вместе составляют примерно 75% от общего мышечного коллагена (McKee et al., 2019). Предполагается, что прочные параллельные волокна коллагена I типа, присутствующие в эндо-, пери- и эпимизиуме, придают мышцам прочность на разрыв и жесткость, тогда как коллаген III типа образует рыхлую сеть волокон, которая придает эластичность мышцам. эндо- и перимизий (Кованен, 2002). Коллаген типа IV, спиральная молекула, образует сетчатую структуру, которая составляет основу базальной пластинки (Sanes, 2003).Коллаген типа VI был обнаружен в эпимизиальном, перимизиальном и эндомизиальном интерстиции, но, в частности, в окрестностях базальной мембраны, где он взаимодействует с карбоксиконцевым глобулярным доменом коллагена типа IV (Kuo et al., 1997). Интересно, что коллаген VI обладает нетипичными неколлагеновыми участками, формирующими отчетливую микрофибриллярную сеть в большинстве соединительных тканей (Maaß et al., 2016). Мутации коллагена VI приводят к заболеваниям с сочетанным вовлечением мышц и соединительной ткани, включая врожденную мышечную дистрофию Ульриха, миопатию Бетлема, аутосомно-доминантную мышечную дистрофию пояса конечностей и аутосомно-рецессивный миосклероз (Bushby et al., 2014).
Таблица 1. Обзор коллагеновых компонентов внеклеточного матрикса скелетных мышц.
Коллаген типов XII, XIV, XIX и XXII принадлежит к ассоциированным с фибриллами коллагенам с прерывистыми тройными спиралями (FACIT; Chiquet et al., 2014; Calvo et al., 2020), при этом коллаген типа XXII, по-видимому, экспрессируется исключительно в тканях. соединения, такие как мышечно-сухожильные соединения в скелетных и сердечных мышцах (Koch et al., 2004).
Биоинформатические инструменты для скрининга протеома человека нормальных и больных тканей позволили охарактеризовать глобальный состав протеома ЕСМ, или «матрисомы.В общей сложности 1027 генов были связаны с ЕСМ, посредством чего основные белки матрисомы (гликопротеины ЕСМ, коллагены и протеогликаны) можно было отличить от белков, связанных с матрисомами (аффилированные с ЕСМ белки, регуляторы ЕСМ и секретируемые факторы, которые могут взаимодействовать с core ECM белки) (Naba et al., 2016). Учитывая сложность ткани скелетных мышц человека, включающей многоядерные мышечные волокна, иммунные клетки, эндотелиальные клетки, мышечные стволовые клетки, немиогенные мезенхимальные предшественники и другие мононуклеарные клетки (Bentzinger et al., 2013a), будущие исследования будут необходимы для выяснения вклада каждой из этих клеток в структуру и реконструкцию IMCT. Сигнатуры генов, полученные, например, из последовательности РНК изолированных мышечных волокон и других типов клеток, представляют собой многообещающий инструмент для деконволюции основной массы скелетной мышечной ткани (Rubenstein et al., 2020).
Физиологическая регуляция генов ECM
Гомеостаз ЕСМ поддерживается посредством точно настроенных анаболических и катаболических процессов, которые регулируются различными факторами роста, протеогликанами и ферментами, ответственными за деградацию коллагена.Было обнаружено, что после связывания с мембранно-связанными рецепторами факторы роста, принадлежащие к суперсемейству трансформирующих факторов роста бета (TGF-β), индуцируют фосфорилирование белков Smad, которые передают внеклеточные сигналы в ядро, где они активируют транскрипцию нижележащих генов, что приводит к продукции коллагена. (Макдональд и Кон, 2012). Другим, хотя и менее описанным, фактором аналогичной функции является фактор роста соединительной ткани (CTGF), избыточная экспрессия которого, как сообщается, провоцирует дистрофический фиброз мышц и функциональные нарушения (Morales et al., 2011).
Функция этих анаболических факторов в основном регулируется небольшими протеогликанами, богатыми лейцином (SLRP). Декорин, прототип этого семейства, деактивирует профибротический TGF-β и CTGF (Zhu et al., 2007; Brandan and Gutierrez, 2013), а также ограничивает фибриллогенез путем прямого связывания с коллагеном I типа (Reese et al., 2013) . Другой SLRP представлен бигликаном, который конкурирует с декорином за тот же сайт связывания на коллагене (Schönherr et al., 1995) и, вероятно, играет роль как в формировании, так и в регенерации мышц (Brandan et al., 2008).
Транскрипционная регуляция образования белков, по-видимому, является важным фактором пластичности ECM. В этом отношении было показано, что экспрессия белка в скелетных мышцах слабо регулируется на уровне мРНК, что приводит к большим различиям в содержании мРНК и белка в различных тканях (Wang et al., 2019). Интересно, что характер регуляции белка зависит от функции белка, при этом ассоциация между мРНК и белком выше для ECM и организации коллагеновых фибрилл (Makhnovskii et al., 2020). Другим интересным аспектом регуляции количества белков ЕСМ является тот факт, что индукция транскрипции для коллагена, по-видимому, довольно медленная, поскольку для полной индукции транскрипции требуется почти 3 дня. Напротив, скорость секреции адаптируется быстро, поскольку она повышается менее чем за 1 час. В клетках с высоким уровнем продуцирования коллагена этот путь контролируется посттранскрипционной регуляцией, которая требует контроля обратной связи между скоростью секреции и трансляции (обзор у Schwarz, 2015).
Что касается ремоделирования ткани, два семейства ферментов, матриксные металлопротеиназы (MMP) и тканевые ингибиторы металлопротеиназ (TIMP), участвуют в регуляции гомеостаза ECM. ММП представляют собой протеолитические ферменты, которые разрушают различные типы коллагенов и ингибируются ТИМП (Visse and Nagase, 2003; Alameddine, 2012). В частности, MMP-1 и MMP-8 инициируют деградацию коллагенов I и III (преобладающих в эндо-, пери- и эпимизиуме), тогда как MMP-2 и MMP-9 расщепляют коллаген IV типа (основной коллагеновый компонент базальная мембрана) (Corcoran et al., 1996). ТИМП-1, -2 и -4 способны ингибировать все известные ММП (Christensen and Purslow, 2016).
ECM и сила скелетных мышц
Взаимодействие актина и миозина, а также многих других саркомерных белков приводит к укорочению мышечных волокон. Традиционные биомеханические модели часто изображают мышечно-сухожильные единицы как системы, в которых силы, возникающие в результате укорочения волокна, передаются продольно вдоль мышечного волокна и далее, в мышечно-сухожильном соединении, на сухожилие.Рядом с мышечно-сухожильным соединением миофибры имеют отростки, похожие на пальцы, которые образованы из инвагинаций плазматической мембраны (Knudsen et al., 2015). Эта структура увеличивает площадь поверхности, доступную для передачи усилия. Ожидается, что передача силы будет происходить между пальцевидными отростками мышечного волокна и коллагеновыми волокнами, расположенными внутри инвагинаций, посредством срезания базальной пластинки (Huijing, 1999). Содержащиеся здесь коллагены относятся к типу XXII, который образует внутренний слой, а также к коллагенам III, VI, XII и XIV, которые расположены дальше от мембраны мышечных волокон (Jakobsen et al., 2017). Хотя его точная роль до сих пор неясна, интересно отметить, что в мышцах коллаген XXII располагается исключительно в мышечно-сухожильном соединении. У рыб-зебр дефицит коллагена XXII, как было обнаружено, приводит к мышечной дистрофии (Charvet et al., 2013), предполагая, что этот коллаген может служить для поддержания структурной целостности и стабилизации мышечно-сухожильного соединения.
Учитывая тот факт, что значительная часть волокон в длинных мышцах оканчивается внутрипучкообразно, не достигая непосредственно сухожилия (Barrett, 1962; Hijikata et al., 1993), однако ясно, что мышечно-сухожильный путь не может представлять собой единственный механизм передачи силы. Внутрисосудистые концевые волокна должны полагаться на среду, расположенную параллельно им, чтобы передавать свои силы на пассивные компоненты опорно-двигательной системы (Sheard, 2000). Как впервые было обнаружено Street (1983), именно сеть IMCT в ECM способствует такой боковой передаче сократительной силы. Передача силы через сеть IMCT происходит от сократительных белков через костамеры к эндомизию (Bloch and Gonzalez-Serratos, 2003; Peter et al., 2011) — как показывают исследования моделирования, через срезание (Sharafi and Blemker, 2011; Zhang and Gao, 2012) — и далее в перимизий, который окончательно сливается с апоневрозами и сухожилиями (Passerieux et al., 2007). Первые сведения о пропорциях передачи продольной и поперечной силы в поперечнополосатых мышцах получены из элегантных экспериментов Huijing et al. (1998). После разрыва прямых соединений нескольких головок длинного разгибателя пальцев рук крысы, соответствующих 55% общей мышечной массы, от суставного сухожилия, Huijing et al.(1998) наблюдали, что сила поддерживалась на уровне 84% от силы неповрежденной мышцы. Совсем недавно Ramaswamy et al. (2011) использовали коромысло для прямого измерения сил, передаваемых по продольным и поперечным путям. Их результаты не только подтвердили, что более 50% силы передавалось в боковом направлении, но также показали, что передача боковой силы была значительно снижена как у дистрофических, так и у старых грызунов. Позже их результаты были подтверждены Чжаном и Гао (2014).
Некоторые аргументы предполагают, что латеральная передача силы является биомеханической необходимостью для поддержания целостности мышц и повышения эффективности сокращения.Во-первых, он помогает распределять сократительные силы по всей поверхности миофибрилл, что снижает механическое напряжение и защищает волокна от чрезмерного растяжения. Это может быть особенно важно в концевых участках волокна, которые обычно сужаются и поэтому не подходят для выдерживания чрезмерных усилий (Monti et al., 1999). Косвенным подтверждением этой гипотезы являются исследования с участием пожилых людей (Hughes et al., 2016) или пациентов, страдающих дистрофией Дюшенна (Virgilio et al., 2015), у которых дистрофин (i.е. костамерный белок, который устанавливает механическую связь между цитоскелетом, сарколеммой и ВКМ и, таким образом, способствует передаче латеральной силы), либо теряется, либо нарушается, и восприимчивость к травмам от растяжения мышц увеличивается.
Кроме того, считается, что передача латеральной силы приводит к сокращению волокон моста либо в разное время, либо в разной степени (Yucesoy et al., 2006), что помогает поддерживать выравнивание волокон и, таким образом, структурную целостность мышцы (Purslow, 2002).Недавно Dieterich et al. (2017) сравнили начало сокращения, определенное электромиографией и УЗИ в M-режиме. Как ни странно, авторы обнаружили, что начало движения предшествует электромиографическому сигналу в ~ 20% исследований, что можно объяснить переносом боковой силы. Действительно, хотя продольная передача сил может быть задержана из-за необходимости натягивать эластичные элементы, размещенные последовательно с мышцей (Nordez et al., 2009), трансламинарное сцепление сдвига между мышечными волокнами и сетью IMCT может обеспечить немедленную передачу силы. .Наконец, передача боковой силы обеспечивает механизм, посредством которого сила может по-прежнему генерироваться и передаваться от мышечных волокон, которые прерваны из-за микротравм или во время роста мышц (Purslow, 2010).
Помимо своей роли в латеральной передаче сократительной силы, ECM может также влиять на укорочение мышечных волокон. Часто предполагается, что сократимость миофибрилл ограничивается геометрией составляющих его саркомеров: саркомера и, таким образом, укорачивание волокна прекращается, когда полосы z входят в контакт с миозиновыми нитями.Однако эти идеи рассматривают только поведение саркомера как независимого исполнительного механизма. В условиях in vivo мышечные волокна встраиваются в сеть IMCT, что может препятствовать укорочению волокон. Действительно, принцип постоянного объема (Baskin and Paolini, 1967) диктует, что во время укорачивания мышечные волокна должны подвергаться радиальному расширению, что уже давно экспериментально подтверждено даже на саркомерном уровне (Brandt et al., 1967). Новые вычислительные модели и измерений in situ в мышцах лягушки Азизи и др.(2017) продемонстрировали, что сокращение мышц затрудняется, когда радиальное расширение ограничивается физическими ограничениями. Следовательно, изменения количества и механических свойств сети IMCT, в которую встроены мышечные волокна, могут напрямую влиять на сократимость скелетных мышц. Такой сценарий может быть представлен мышечным фиброзом (Gillies et al., 2017).
ECM в развитии, росте и восстановлении скелетных мышц
Помимо передачи силы, ECM скелетных мышц выполняет несколько важных функциональных ролей.Очевидно, сеть ММКИ обеспечивает механическую поддержку мышечным волокнам, а также поддерживающим их нервам и кровеносным сосудам. Кровеносные капилляры проходят в промежутках, занятых эндомизием, их количество и плотность зависят от размера мышечных волокон (Janácek et al., 2009). Помимо этой наиболее очевидной роли, взаимодействие между миобластами, дифференцированными мышечными волокнами и компонентами внеклеточного матрикса имеет центральное значение для эмбриогенного развития, дальнейшего роста и восстановления мышечной ткани.
Клеточным источником коллагеновых компонентов мышечного ECM являются специальные фибробласты IMCT, которые происходят из различных эмбриогенных источников, включая сомиты (Nowicki et al., 2003), мезодерму латеральной пластинки (Pearse et al., 2007) и нервную систему. клетки гребня (Olsson et al., 2001). Поскольку они продуцируют не только фибробласты, но и адипогенные клетки, фибробласты IMCT могут рассматриваться как фиброипогенные предшественники (Uezumi et al., 2010). Недавние исследования предоставили доказательства того, что, помимо этих очевидных ролей, фибробласты IMCT и производимые ими соединительные ткани влияют на миогенез (т.е.д., образование мышечных предшественников и их дифференциация в многоядерные миофибриллы) и морфогенез мышц (то есть процесс, в котором миофибры собираются в мышцы), таким образом действуя как важные регуляторы развития мышц. Эти сложные регуляторные процессы, происходящие во время эмбриогенного развития, здесь подробно не рассматриваются, но были подробно рассмотрены в других источниках (Nassari et al., 2017; Sefton and Kardon, 2019). Вкратце, IMCT направляет мышечных предшественников к их назначенным областям-мишеням посредством комбинации привлекательных (фактор роста гепатоцитов, фактор, производный стромальными клетками) и отталкивающих сигналов (эфрин) (Dietrich et al., 1999; Swartz et al., 2001). Посредством множества факторов транскрипции, экспрессируемых в фибробластах IMCT, IMCT затем способствует пролиферации, выживанию и дифференцировке соседних миобластов в зрелые миофибриллы (Kardon et al., 2003; Hasson et al., 2010; Iwata et al., 2013; Vallecillo -García et al., 2017). Таким образом, можно предположить, что IMCT служит мезодермальным препаттерном, который контролирует участки дифференцировки миофибрилл и, следовательно, окончательное положение, размер и форму мышц.
Как постмитотические ткани, скелетные мышцы зависят от сателлитных клеток для адаптации и регенерации на протяжении всей жизни. Эти стволовые клетки располагаются в специализированных нишах между сарколеммой мышечных волокон и их инкапсулирующими базальными мембранами. Поддержание, активация и дифференцировка сателлитных клеток регулируются сложными каскадами факторов транскрипции. Для подробного обзора этих сотовых схем читатели могут обратиться к недавнему обзору Almada and Wagers (2016). Имея особое отношение к этой рукописи, растущее количество доказательств указывает на то, что судьба сателлитных клеток также сильно зависит от взаимодействий с нишей ECM, в которой они находятся.Действительно, как динамическая среда ниша стволовых клеток передает механические и химические сигналы, которые действуют, защищая покоящиеся стволовые клетки или индуцируя активацию, пролиферацию и дифференцировку.
В состоянии покоя сателлитные клетки экспрессируют канонический клеточный регулятор спаренный бокс-белок 7 (PAX7) (Olguin and Olwin, 2004). Исследования in vitro показали, что большая часть сателлитных клеток экспрессирует PAX7 при культивировании на матригеле, смеси белков ЕСМ и факторов роста (Wilschut et al., 2010; Грефте и др., 2012). Дальнейшее подтверждение идеи, что ECM активно участвует в поддержании покоя сателлитных клеток, происходит из сообщений о том, что сателлитные клетки, удаленные из своей ниши, быстро входят в клеточный цикл и теряют свою способность к миогенной дифференцировке (Gilbert et al., 2010). Интересно, что сателлитные клетки, по-видимому, также способны воспринимать различные механические свойства ECM и реагировать на них. Фактически, экспрессия PAX7 и выживаемость сателлитных клеток выше при культивировании на гидрогелях, имитирующих физиологическую жесткость мышц (Gilbert et al., 2010). Кроме того, сателлитные клетки, культивируемые на мягком гидрогеле, обладают большей функциональной способностью после трансплантации в мышцу реципиента (Cosgrove et al., 2014).
Кроме того, было показано, что компоненты ECM влияют на деление стволовых клеток. В частности, белки фибронектин (Bentzinger et al., 2013b) и коллаген VI (Urciuolo et al., 2013), а также протеогликаны синдекан 3, синдекан 4, перлекан и декорин (Cornelison et al., 2001; Brack et al. ., 2008) были идентифицированы как составляющие ниши, влияющие на баланс между дифференцировкой и самообновлением и, таким образом, на поддержание регенеративной способности скелетных мышц.
При мышечной травме или в ответ на повышенную нагрузку обычно в основном покоящиеся сателлитные клетки активируются и дифференцируются в миобласты, чтобы окончательно слиться в зрелые миофибриллы. Хотя этот процесс требует своевременной экспрессии различных факторов транскрипции, таких как миогенный фактор 5, белок миогенной детерминации или миогенин (Almada and Wagers, 2016), несколько исследований указывают на влияние ECM на каждый из этих этапов. Эксперименты с миобластами мышей (Grefte et al., 2012) или свиней (Wilschut et al., 2010) показали, что на слияние миобластов положительно влияет матригель, но не отдельные субстраты, присутствующие в нише ECM. Вклад отдельных белков все еще плохо изучен, однако сопутствующее присутствие поли- D -лизина и ламинина (Boonen et al., 2009), гликозаминогликанов (Rønning et al., 2013) и протеогликанов гепаринсульфата (Gutiérrez и Brandan, 2010), по-видимому, играют важную роль в пролиферации и дифференцировке сателлитных клеток. После активации стволовых клеток скелетных мышц локальное ремоделирование ВКМ сопровождается отложением ламинина-α1 и ламинина-α5 в базальной пластинке ниши сателлитных клеток (Rayagiri et al., 2018). На мышах было показано, что мышечные сателлитные клетки продуцируют коллагены ЕСМ для поддержания покоя клеточно-автономным образом, при этом коллаген V является критическим компонентом покоящейся ниши, поскольку истощение приводит к аномальному вхождению в клеточный цикл и постепенному уменьшению стволовых клеток. бассейн (Багдади и др., 2018). Так же, как для поддержания покоя, адекватные механические свойства ниши ECM также могут быть важны для созревания сателлитных клеток. Действительно, было обнаружено, что миотрубки оптимально дифференцируются на субстратах с мышечной жесткостью (Engler et al., 2004). В совокупности эти данные предполагают, что усиление жесткости ВКМ, сопровождающее как различные скелетно-мышечные нарушения, так и процесс старения, может отрицательно влиять на регенеративную способность мышцы.
Ремоделирование мышц ECM со старением
В более старшем возрасте скелетные мышцы обычно демонстрируют фиброзную морфологию (Lieber and Ward, 2013). В отличие от фасциального уплотнения, при котором может сохраняться общая структура коллагенов (Pavan et al., 2014), возрастной мышечный фиброз характеризуется потерей четкой двунаправленной ориентации решетки здоровых перимизиальных волокон коллагена и его замещением. из-за неустойчивой оптоволоконной сети со сниженным образованием складок (Ярвинен и др., 2002). Кроме того, может увеличиваться абсолютное содержание коллагена и (неферментативное) сшивание коллагеновых волокон (Haus et al., 2007b). Таким образом, модуль упругости ЕСМ может быть увеличен примерно в 35 раз (с ~ 12 кПа в молодых до ~ 418 кПа в старых мышцах; Yin et al., 2013), причем этот эффект обусловлен накоплением плотно упакованных и сильно сшитый коллаген (Wood et al., 2014). У крупных и долгоживущих животных, таких как тюлени Уэдделла, наблюдалось увеличение внеклеточного пространства на 35-40% по мере увеличения общего и относительного содержания коллагена с возрастом.Однако это увеличение связано со сдвигом в сторону более высокого отношения коллагена типа I к коллагену типа III (Hindle et al., 2009). Более того, концентрация коллагена IV типа повышается в базальной пластинке медленно сокращающихся мышц, тогда как концентрация ламинина, по-видимому, снижается с возрастом (Kovanen et al., 1988). Было также показано, что повышенное отложение белков базальной пластинки вытесняет сателлитные клетки из их ниш, что влияет на регуляцию делений сателлитных клеток (Snow, 1977) и может объяснять меньшее количество сателлитных клеток, обычно учитываемых в старых мышцах по сравнению с молодыми мышцами. (Brack et al., 2007). Утрата и функциональная инактивация стволовых клеток, которые негативно влияют на гомеостаз тканей, могут рассматриваться как общий признак старения (López-Otín et al., 2013), который следует рассматривать как универсальную силу, приводящую к старению мышц (Brack and Muñoz-Cánoves, 2016) и другие ткани (Oh et al., 2014). Помимо воздействия на сателлитные клетки, нерегулируемая базальная пластинка также, как ожидается, нарушит регенеративную способность мышц из-за неадекватной поддержки мышечных волокон и дезорганизованной ориентации каркаса (Sanes, 2003).Обзор, включающий обширный обзор эффектов старения на ECM скелетных мышц, был недавно опубликован Etienne et al. (2020).
Интересно, что данные транскрипционного профилирования мышц молодых и старых крыс предполагают, что из 682 наборов зондов, которые значительно различались между молодыми и старыми животными, 347 генов фактически уменьшились (а не увеличились) в старых / саркопенических мышцах по сравнению с молодыми мышцами. Было показано, что из этих генов 24% играют биологическую роль в ECM и клеточной адгезии (Pattison et al., 2003). Эти данные подтверждают гипотезу, что возрастные изменения в ECM могут быть вызваны сниженной способностью к деградации, а не повышенным синтезом коллагеновых структур. В частности, ММП, по-видимому, играют важную роль в этих процессах (de Sousa Neto et al., 2018). Это также подтверждается данными, которые предполагают снижение способности структур ВКМ к ремоделированию структур ВКМ в старых мышцах, вызванное упражнениями с отягощениями (Wessner et al., 2019). Хотя механизмы еще полностью не изучены, также считается, что эти изменения напрямую ухудшают функцию мышц, препятствуя сократимости волокон (Azizi et al., 2017) и передача поперечной силы (Sharafi and Blemker, 2011).
Нарушение регуляции ECM скелетных мышц вследствие метаболических нарушений
Хорошо известно, что скелетные мышцы играют важную роль в стимулированном инсулином поглощении глюкозы (Richter and Hargreaves, 2013). Роль ECM в этом контексте может быть менее ясной. Мышцы-специфичные интегрин-β1-дефицитные мыши демонстрируют снижение скорости инфузии глюкозы, стимулированной инсулином, и клиренса глюкозы, несмотря на отсутствие изменений в приеме пищи, весе, глюкозе натощак, уровнях инсулина и экспрессии белка GLUT4 (Zong et al., 2009), подтверждая связь между аберрантной передачей сигналов интегрина и развитием диабета 2 типа. Более того, на животной модели диабета было показано, что нарушение чувствительности к инсулину связано со снижением уровней белка изоформы дистрофина Dp427 и субкомплекса альфа / бета-дистрогликана (Mulvey et al., 2005).
Повышенное количество коллагена типа I и III было обнаружено как у пациентов с диабетом 2 типа, так и у недиабетиков с ожирением (Berria et al., 2006), а перекорм у людей был связан с увеличением экспрессии генов, связанных с IMCT (коллагены I, III, IV, V, SPARC, интегрин; Tam et al., 2014) и изменения генных путей, связанные с взаимодействием рецепторов ECM, фокальной адгезией и сращиванием (Tam et al., 2017). Однако кормление мышей диетой с высоким содержанием жиров привело к снижению уровней экспрессии генов COL1, COL3 и COL6, но не уровня белка (Tam et al., 2015).
Деградация коллагенов посредством MMPs, как было показано, является важным компонентом ремоделирования ECM (Cui et al., 2017). Может ли это быть правдой и в контексте диабета, было исследовано в исследовании на животных.Интересно, что генетическое истощение MMP9 не вызывало инсулинорезистентности у худых мышей, несмотря на то, что приводило к увеличению коллагена IV. Однако, когда мышей кормили диетой с высоким содержанием жиров, делеция вызывала глубокое состояние инсулинорезистентности. Эти результаты еще больше усиливают роль компонентов ММКИ в развитии мышечной инсулинорезистентности, особенно в состоянии переедания (Kang et al., 2014).
Наконец, гиалуронан, основной компонент внеклеточного матрикса, увеличивается при ожирении у мышей, вызванном диетой с высоким содержанием жиров.Для лечения инсулинорезистентности с сопутствующим уменьшением жировой массы, размера адипоцитов, а также печеночной и мышечной инсулинорезистентности (Kang et al., 2013) предлагается лечение с помощью PEGPh30, который дозозависимо снижает гиалуронан в мышечном ECM.
Подводя итог, можно сказать, что различные компоненты ECM подвержены влиянию на различных стадиях диабета. Исследования о том, связан ли диабет со слабостью мышц, неоднозначны (Leong et al., 2015; Li et al., 2016), и еще предстоит выяснить, имеют ли изменения в путях, связанных с ECM, непосредственное участие в этом контексте.
Адаптация к физическому воспитанию и неиспользование
Первые свидетельства, указывающие на гибкость ММКИ в ответ на физическую активность, были опубликованы еще в 1970-х годах, когда Suominen и Heikkinen (1975) и Suominen et al. (1977) обнаружили более высокие уровни пролилгидроксилазы (фермента, способствующего биосинтезу коллагена) у тренированных на выносливость спортсменов, а также в продольном исследовании после восьми недель аэробных тренировок. Позже было обнаружено, что влияние упражнений на выносливость на активность проколлагеновых ферментов более выражено в красных мышцах по сравнению с белыми мышцами (Takala et al., 1983). Прямые измерения содержания коллагена, впервые проведенные в конце 1980-х годов, подтвердили, что содержание коллагена (тип IV) увеличилось в устойчивой к усталости камбаловидной мышце крыс после пожизненных тренировок на выносливость (Kovanen et al., 1988). Несмотря на вызванное физическими упражнениями увеличение коллагена, Gosselin et al. (1998) обнаружили, что жесткости мышц, наблюдаемой с возрастом, можно противодействовать с помощью упражнений на выносливость, которые авторы связывают с уменьшением сшивания гидроксилизилпиридинолином коллагеновых волокон.
Эффекты иммобилизации на ECM скелетных мышц не совсем однозначны. Ранние исследования Karpakka et al. (1990, 1991) обнаружили, что у крыс снижается как активность гидроксилазы, так и содержание гидроксипролина (аминокислоты, составляющей коллагены). Позже было обнаружено, что изменения содержания коллагена в ответ на кратковременную иммобилизацию или неиспользование были довольно небольшими (Savolainen et al., 1988; Haus et al., 2007a), что можно объяснить относительно низкой скоростью обновления. В более позднем исследовании, напротив, было обнаружено, что содержание коллагена I и биомеханические свойства (модуль упругости, максимальное напряжение и предел текучести) фасции голени, покрывающей трехглавую мышцу surae крысы, значительно увеличиваются после всего лишь 21 дня разгрузки задней конечности. (Хуанг и др., 2018). Интересно, что эти изменения можно предотвратить, применяя вибрацию к задним лапам крыс дважды в день. У людей, не занимающихся спортом, иммуногистохимическое окрашивание не показало изменений в плотности сети коллагена I после 60 дней постельного режима. Напротив, у субъектов, выполняющих протокол контрмер, состоящий из реактивных прыжков на санях, иммунореактивность коллагена I была снижена по сравнению с исходными уровнями (Schoenrock et al., 2018).
Еще одна модель, которая позволяет изучить адаптивность ECM мышц, — это функциональная перегрузка, вызванная устранением хирургического синергиста.В одном из первых соответствующих исследований Уильямс и Голдспинк (1981) перерезали сухожилия подошвенных и икроножных мышц самцов крыс, чтобы перегрузить камбаловидные мышцы. Гипертрофия мышц, наблюдаемая через три недели после тенотомии, сопровождалась увеличением концентрации IMCT (> 45%) и отношения IMCT к мышечной ткани. Гистологический анализ также показал, что увеличение IMCT в основном связано с утолщением эндомизия. Сосредоточившись на мышечно-сухожильном соединении, Zamora и Marini (1988) провели аналогичные эксперименты и изолировали подошвенную мышцу крысы с помощью тенотомии камбаловидной мышцы и удаления икроножных мышц.По сравнению с контрольными животными фибробласты, расположенные в мышечно-сухожильном соединении, развивали более высокую степень активации цитоплазмы, ядра и ядрышка уже после одной-двух недель функциональной перегрузки. В более недавнем исследовании было проверено влияние ИЛ-6 на ремоделирование ВКМ, вызванное перегрузкой, путем сравнения мышей дикого типа и мышей с нокаутом по ИЛ-6 (White et al., 2009). В то время как увеличение площади поперечного сечения миофибрилл было одинаковым после 21 дня функциональной перегрузки, увеличение сырого веса мышц было значительно больше у мышей с нокаутом по IL-6.Гистологический анализ подтвердил, что этот избыточный прирост мышечной массы можно объяснить значительно большим увеличением содержания несократительной ткани и концентрации гидроксипролина, что свидетельствует о содержании коллагена и фиброзе. В соответствии с этим наблюдением, уровни мРНК проколлагена-1, IGF-1 и TGF-β были значительно выше у мышей с дефицитом IL-6. Напротив, экспрессия мРНК MyoD, фактора транскрипции, необходимого для мио-, а не фиброгенной дифференцировки сателлитных клеток (Zammit, 2017), была значительно ослаблена у животных, лишенных IL-6.В совокупности эти результаты показывают, что устранение синергиста вызывает увеличение содержания IMCT и, в частности, утолщение эндомизиальных структур в перегруженных мышцах. Эти адаптации могут служить для модуляции несокращающихся структур мышц в соответствии с повышенными функциональными требованиями. IGF-1, по-видимому, играет важную роль в регуляции этого процесса, поскольку было показано, что недостаток IGF-1 приводит к чрезмерному накоплению IMCT и, возможно, к нарушению регенеративного потенциала мышц.
Одно из первых исследований по тестированию и сравнению различных форм упражнений с отягощениями у мужчин было проведено Brown et al.(1999), которые сообщили, что после одного приступа концентрических сокращений маркеры распада коллагена (гидроксипролин и коллаген сывороточного типа I) не увеличивались. Напротив, эксцентрическое действие мышц увеличивало уровень коллагена в сыворотке более чем на 40% в течение 9 дней после тренировки, что указывает на то, что эксцентрические сокращения могут быть более эффективными в стимулировании распада коллагена. Эти результаты были подтверждены в двух более поздних исследованиях, аналогичным образом использовавшихся с использованием высокоинтенсивных эксцентрических упражнений, которые показали как повышенный процессинг проколлагена и содержание коллагена типа IV, так и более высокую активность ММП и ТИМП (Crameri et al., 2004; Mackey et al., 2004). Интересно, что Crameri et al. (2004, 2007) также сообщили об увеличении тенасцина С, гликопротеина, присутствующего в ЕСМ, который, как предполагается, направляет миграцию клеток после повреждения, независимо от того, было ли повреждение мышц вызвано произвольным или электрически индуцированным повреждением мышц. Временное повышение активности тенасцина С и других гликопротеинов ЕСМ (например, фибронектина и гиалуроновой кислоты) обычно называют «переходной матрицей», появление которой считается важным первым шагом для успешного восстановления мышц, так как дает важные сигналы, управляющие регенеративный потенциал мышечных стволовых клеток (Calve et al., 2010; Tierney et al., 2016). Сообщается, что высвобождение гликопротеинов ECM сопровождается повышенной активностью MMP-9 у молодых, но снижением активности MMP-9 и MMP-15 у пожилых людей (Wessner et al., 2019). Эти данные свидетельствуют о том, что острые упражнения с отягощениями вызывают катаболический ответ в молодых мышцах, но этот эффект может быть нарушен в более старшем возрасте. Последующая анаболическая реакция, характеризующаяся повышенной регуляцией структурных коллагенов (I, III, IV) и ламинина, как было обнаружено, происходит со значительной задержкой, что позволяет предположить, что восстановление мышц, вызванное острым приступом повреждающих (удлинительных) сокращений, следует за двухфазный характер (Mackey et al., 2011; Hyldahl et al., 2015). Интересно, что недавнее исследование Sorensen et al. (2018) обнаружили, что внешний вид переходной матрицы у физически активных пожилых людей притупляется по сравнению с молодыми людьми. Это наблюдение поддерживает представление о том, что дисрегулируемые сигналы ECM могут быть ответственны за увеличение отложения ECM и снижение активности стволовых клеток, обычно наблюдаемых в более старых мышцах (Grounds, 1998).
Одно из первых исследований по прямому сравнению различных форм мышечного сокращения с точки зрения их острого ремоделирующего потенциала ВКМ было опубликовано Heinemeier et al.(2007). Эти авторы провели исследование на грызунах и обнаружили, что активность генов, связанных с биосинтезом коллагена (например, коллагенов I и III), а также факторов роста (например, TGF-β1) повышается после всех форм физических упражнений, но в большей степени это происходит. после эксцентрической тренировки. У людей также было предложено увеличить скорость фракционного синтеза белка коллагена после острой эксцентрической тренировки по сравнению с концентрической тренировкой (Holm et al., 2017), хотя это понятие не бесспорно (Moore et al., 2005). В совокупности эти результаты предполагают, что особенно эксцентрические упражнения являются мощным стимулом, который вызывает микротравмы и обмен клеток IMCT, причем последний, как предполагается, представляет собой попытку организма предотвратить повторное повреждение мышцы (Mackey et al., 2011; Hyldahl et al. , 2015; Takagi et al., 2016). Фактически, снижение активности ММП после продолжительной тренировки, состоящей из электрически вызванных изометрических сокращений у крыс, может отражать успешное усиление ВКМ (Ogasawara et al., 2014), тогда как продолжительное повышение активности ММП и ТИМП в подошвенной мышце мышей после хирургического удаления икроножной и камбаловидной мышц может указывать на продолжающееся ремоделирование ВКМ (Mendias et al., 2017).
Помимо режима сокращения, ECM скелетных мышц также может быть чувствительным к интенсивности упражнений. Carmeli et al. (2005) протестировали влияние бега на беговой дорожке с высокой или низкой интенсивностью на крысах и обнаружили, что MMP-2 (один из ферментов, ответственных за расщепление коллагена IV, в основном присутствующего в базальной мембране мышц) повышается после упражнений высокой интенсивности. Только. На людях, напротив, одно исследование Holm et al. (2010) сравнили эффекты односторонних упражнений на разгибание колен, выполняемых с низкой или высокой (16% или 70% от индивидуального максимума одного повторения, соответственно) интенсивностью, с количеством повторений, скорректированным в соответствии с вмешательствами для общей нагрузки. поднял.В этом исследовании скорость фракционного синтеза коллагена была равномерно увеличена после обоих вмешательств.
Что касается адаптации ЕСМ к длительным тренировкам с отягощениями, существуют только данные исследований на животных. де Соуза Нето и др. (2018) сообщили, что 12 недель тренировок с отягощениями, состоящих из подъемов по лестнице с прогрессивными дополнительными нагрузками, эквивалентными 65–100% максимальной несущей способности каждого индивидуума, активировали активность ММП-2 в подошвенных мышцах старых крыс, в то же время подавляя ММП-2. 2 и ММП-9 в кровообращении.Вывод авторов о том, что тренировки с отягощениями могут, таким образом, быть полезным инструментом для поддержания ремоделирования ВКМ в более старшем возрасте, недавно получил эмпирическую поддержку в другом исследовании обучения на крысах, которое использовало тот же протокол обучения и показало пониженное отложение соединительной ткани у тренированных пожилых людей. мышцы (Guzzoni et al., 2018).
Подводя итог, можно сказать, что несколько исследований, посвященных изучению острых эффектов физической активности как у грызунов, так и у мужчин, показали, что упражнения могут стимулировать как деградацию, так и синтез коллагена в скелетных мышцах.Восстановление микротравм, вызванных физической нагрузкой, следует двухфазному паттерну, при котором гликопротеины сначала создают переходную матрицу для управления катаболическими процессами, а анаболические процессы, укрепляющие структуру IMCT, происходят со значительной задержкой. Потенциал упражнений для ремоделирования ВКМ, по-видимому, зависит от режима сокращения, при этом эксцентрические сокращения вызывают большую реакцию, чем концентрическое или изометрическое мышечное действие. Доступно немного исследований, в которых проверяются результаты упражнений с различной интенсивностью, и пока результаты позволяют предположить, что распад белка (но не синтез) может быть сильнее спровоцирован более высокой интенсивностью.Неиспользование резко снижает активность ферментов, связанных с биосинтезом коллагенов, хотя на уровне белка изменения происходят с медленной скоростью. Поперечные сравнения с участием (в основном тренированных на выносливость) грызунов показывают, что хроническая физическая активность может привести к усилению фенотипа ММКИ. Единственные доступные на сегодняшний день долгосрочные долгосрочные обучающие исследования были выполнены на грызунах и предполагают, что длительные тренировки с отягощениями могут быть полезны для противодействия чрезмерному накоплению ММКИ в более старшем возрасте.Физиологические и функциональные последствия ремоделирования ММКИ, вызванного тренировкой, требуют дальнейшего изучения.
Заключение
Целью настоящего обзора является обзор современного состояния знаний о ECM скелетных мышц, который играет важную, хотя и часто недооцениваемую роль в поддержании мышечного гомеостаза, влияет на мышечную функцию и адаптацию и может быть ключом к лечению. мышечных и метаболических нарушений, вызванных старением или болезнью.
Как сложная сеть различных коллагенов, гликопротеинов, протеогликанов и эластина, ECM включает сократительные мышечные волокна и через интегрины и гликопротеиновый комплекс, связанный с дистрофином, служит биохимическим и механическим интерфейсом между мышечными клетками и их окружением. Сборке его коллагенового каркаса в основном способствуют факторы роста TGF-β и CTGF, которые регулируются различными протеогликанами, такими как декорин и бигликан. Более того, протеолитические ферменты (MMP), а также их ингибиторы (TIMP) участвуют в регуляции ECM.
Функционально ECM служит средой для передачи сократительной силы, которая может служить не только для повышения эффективности мышечного сокращения, но также для защиты мышечных волокон от чрезмерного напряжения и облегчения заживления микротравм. Помимо своей функциональной роли, ECM активно участвует в регуляции пула сателлитных клеток в мышцах. Ниши ВКМ, образованные между сарколеммой и базальной мембраной, защищают сателлитные клетки от входа в клеточный цикл и, таким образом, помогают поддерживать регенеративный потенциал мышц.Конкретные компоненты ЕСМ, такие как фибронектин, коллаген VI и различные протеогликаны, могут дополнительно способствовать делению стволовых клеток. Напротив, ламинин, гликозаминогликаны и другие протеогликаны, как было показано, способствуют дифференцировке сателлитных клеток и их слиянию в зрелые миофибриллы.
Научные данные также демонстрируют, что ECM скелетных мышц является податливой тканью, которая может подвергаться процессам ремоделирования в результате старения, болезней, физических тренировок или неиспользования.В частности, старение обычно приводит к общему увеличению отложения коллагеновой ткани, изменениям в составе коллагена (переход к более высокому типу коллагена I к типу III) и увеличению неферментативного сшивания коллагена (за счет конечных продуктов гликирования). Эти изменения, которые, возможно, опосредованы снижением активности ММП, приводят к усилению жесткости ECM мышцы и могут нарушать функцию и регенеративный потенциал мышцы.
Ремоделирование внеклеточного матрикса также может быть связано с метаболическими нарушениями, такими как диабет.Было обнаружено, что чрезмерное потребление пищи приводит к повышенной экспрессии генов, связанных с ECM (коллагены I, III, IV, V, SPARC, интегрин). В свою очередь, такое ремоделирование может нарушать передачу сигналов интегрина, тем самым снижая чувствительность к инсулину. Другими компонентами ECM, потенциально представляющими мишени для инсулинорезистентности, являются гиалуронан, комплекс дистрофин-дистрогликан, а также MMP9.
Наконец, ремоделирование ECM может быть вызвано физическими упражнениями. Хотя фактических исследований тренировок немного, есть доказательства того, что упражнения могут резко способствовать как увеличению синтеза коллагена (коллагены I, III, TGF-β1), так и деградации (MMP2, MMP9).Поперечные исследования на людях и продольные исследования на грызунах также предполагают, что такой повышенный обмен коллагена может привести к усилению коллагеновых структур у хронически тренированных субъектов и предотвратить чрезмерное отложение коллагена (то есть фиброз) в мышцах пожилого возраста. Исследования, изучающие последствия длительного неиспользования, показали противоречивые результаты. В то время как в ранних исследованиях сообщалось о снижении активности гидроксилазы и содержания гидроксипролина после кратковременной иммобилизации, в более поздних работах было обнаружено повышение содержания коллагена I после 21 дня разгрузки задних конечностей у крыс, но не изменилось после 60 дней постельного режима у людей.Необходимы дальнейшие исследования и, в частности, обучение людей, чтобы изучить влияние различных методов обучения на структуру и состав ECM.
Авторские взносы
RC участвовал в литературных исследованиях и подготовил рукопись. MG и BW внесли свой вклад в литературные исследования и отредактировали рукопись. Все авторы одобрили окончательную версию рукописи и согласились нести ответственность за все аспекты работы. Все лица, обозначенные как авторы, имеют право на авторство, и все, кто имеет право на авторство, перечислены в списке.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы с благодарностью отмечаем финансовую поддержку этого исследования, полученную от Австрийского научного фонда (FWF): KLI 738-B27.
Список литературы
Aagaard, P., Andersen, J. L., Dyhre-Poulsen, P., Leffers, A.-M., Wagner, A., Магнуссон, С. П. и др. (2001). Механизм увеличения сократительной силы перистых мышц человека в ответ на силовую тренировку: изменения в архитектуре мышц. J. Physiol. 534, 613–623. DOI: 10.1111 / j.1469-7793.2001.t01-1-00613.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Адамс, Г. Р., Хазер, Б. М., Болдуин, К. М., и Дадли, Г. А. (1993). Состав тяжелых цепей миозина скелетных мышц и тренировки с отягощениями. J. Appl. Physiol. 74, 911–915. DOI: 10.1152 / jappl.1993.74.2.911
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Adstrum, S., Hedley, G., Schleip, R., Stecco, C., and Yucesoy, C.A. (2017). Определение фасциальной системы. J. Bodyw. Mov. Ther. 21, 173–177. DOI: 10.1016 / j.jbmt.2016.11.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Алмада, А. Э., и Уэйджерс, А. Дж. (2016). Молекулярная схема судьбы стволовых клеток при регенерации скелетных мышц, старении и болезнях. Нац. Rev. Mol. Cell Biol. 17, 267–279. DOI: 10.1038 / nrm.2016.7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Американский колледж спортивной медицины. (2009). Позиционный стенд Американского колледжа спортивной медицины. Модели прогресса в тренировках с отягощениями для здоровых взрослых. Med. Sci. Спортивные упражнения. 41, 687–708. DOI: 10.1249 / mss.0b013e31810
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Азизи, Э., Деслорье, А.Р., Холт, Н. К., Итон, К. Э. (2017). Устойчивость к радиальному расширению ограничивает мышечное напряжение и работу. Biomech. Модель. Механобиол. 16, 1633–1643. DOI: 10.1007 / s10237-017-0909-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Багдади, М. Б., Кастель, Д., Мачадо, Л., Фукада, С., Бирк, Д. Э., Релэкс, Ф. и др. (2018). Реципрокная передача сигналов с помощью Notch-Collagen V-CALCR удерживает мышечные стволовые клетки в их нише. Природа 557, 714–718. DOI: 10.1038 / s41586-018-0144-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барретт Б. (1962). Длина и способ окончания отдельных мышечных волокон портняжной и задней бедренной мышцы человека. Acta Anat. 48, 242–257. DOI: 10.1159 / 000141843
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бенцингер, К. Ф., Ван, Ю. X., Дюмон, Н. А., и Рудницки, М. А. (2013a). Клеточная динамика в нише мышечных сателлитных клеток. EMBO Rep. 14, 1062–1072. DOI: 10.1038 / embor.2013.182
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бенцингер, К. Ф., Ван, Ю. X., фон Мальцан, Дж., Сулеймани, В. Д., Инь, Х., и Рудницки, М. А. (2013b). Фибронектин регулирует передачу сигналов Wnt7a и рост сателлитных клеток. Стволовые клетки клеток 12, 75–87. DOI: 10.1016 / j.stem.2012.09.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Беррия, Р., Ван, Л., Ричардсон, Д. К., Финлейсон, Дж., Белфорт, Р., Пратипанаватр, Т. и др. (2006). Повышенное содержание коллагена в инсулинорезистентных скелетных мышцах. Am. J. Physiol. Эндокринол. Метаб. 290, E560 – E565. DOI: 10.1152 / ajpendo.00202.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Блох, Р. Дж., И Гонсалес-Серратос, Х. (2003). Передача боковой силы через костамеры в скелетных мышцах. Exerc. Sport Sci. Ред. 31, 73–78. DOI: 10.1097 / 00003677-200304000-00004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боонен, К.Дж. М., Розария-Чак, К. Ю., Баайенс, Ф. П. Т., ван дер Шафт, Д. В. Дж., И Пост, М. Дж. (2009). Важнейшие экологические сигналы из ниши сателлитных клеток: оптимизация пролиферации и дифференцировки. Am. J. Physiol. Cell Physiol. 296, C1338 – C1345. DOI: 10.1152 / ajpcell.00015.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Боппарт, М. Д., и Махмассани, З. С. (2019). Передача сигналов интегрина: связь механической стимуляции с гипертрофией скелетных мышц. Am. J. Physiol. Cell Physiol. 317, C629 – C641. DOI: 10.1152 / ajpcell.00009.2019
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брак А. С., Конбой И. М., Конбой М. Дж., Шен Дж. И Рандо Т. А. (2008). Временное переключение с передачи сигналов notch на Wnt в мышечных стволовых клетках необходимо для нормального миогенеза взрослых. Стволовые клетки клеток 2, 50–59. DOI: 10.1016 / j.stem.2007.10.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брак, А.С., Конбой, М. Дж., Рой, С., Ли, М., Куо, К. Дж., Келлер, К. и др. (2007). Повышенная передача сигналов Wnt во время старения изменяет судьбу мышечных стволовых клеток и увеличивает фиброз. Наука 317, 807–810. DOI: 10.1126 / science.1144090
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brandan, E., Cabello-Verrugio, C., and Vial, C. (2008). Новые механизмы регуляции протеогликанов, декорина и бигликана во время формирования мышц и мышечной дистрофии. Matrix Biol. 27, 700–708. DOI: 10.1016 / j.matbio.2008.07.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Brandt, P. W., Lopez, E., Reuben, J. P., and Grundfest, H. (1967). Взаимосвязь между плотностью упаковки миофиламентов и длиной саркомера в поперечно-полосатой мышце лягушки. J. Cell Biol. 33, 255–263. DOI: 10.1083 / jcb.33.2.255
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Браун С., Дэй С. и Доннелли А. (1999).Косвенные доказательства повреждения скелетных мышц человека и распада коллагена после эксцентрических действий мышц. J. Sports Sci. 17, 397–402. DOI: 10.1080 / 026404199365911
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бушби, К. М., Коллинз, Дж., И Хикс, Д. (2014). «Коллагеновые миопатии типа VI», в Прогресс наследственных заболеваний мягких соединительных тканей, , изд. Дж. Халпер, (Дордрехт: Спрингер), 185–199. DOI: 10.1007 / 978-94-007-7893-1_12
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Calve, S., Одельберг, С. Дж., И Саймон, Х.-Г. (2010). Переходный внеклеточный матрикс определяет поведение клеток во время регенерации мышц. Dev. Биол. 344, 259–271. DOI: 10.1016 / j.ydbio.2010.05.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кальво, А.С., Морено, Л., Морено, Л., Тойвонен, Дж. М., Манзано, Р., Молина, Н. и др. (2020). Коллаген типа XIX: перспективный биомаркер базальных мембран. Neural Regen. Res. 15, 988–995. DOI: 10.4103 / 1673-5374.270299
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кармели, Э., Моас, М., Леннон, С., и Пауэрс, С. К. (2005). Упражнения высокой интенсивности увеличивают экспрессию матричных металлопротеиназ в быстрых волокнах скелетных мышц: упражнения и ММП в быстрых волокнах скелета. Exp. Physiol. 90, 613–619. DOI: 10.1113 / expphysiol.2004.029462
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарве, Б., Гиро, А., Malbouyres, M., Zwolanek, D., Guillon, E., Bretaud, S., et al. (2013). Нокдаун гена col22a1 у рыбок данио вызывает мышечную дистрофию за счет нарушения мышечно-сухожильного соединения. Dev. Camb. Англ. 140, 4602–4613. DOI: 10.1242 / dev.096024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чике М., Бирк Д. Э., Беннеманн К. Г. и Кох М. (2014). Коллаген XII: защита целостности костей и мышц за счет организации фибрилл коллагена. Внутр. Дж.Биохим. Cell Biol. 53, 51–54. DOI: 10.1016 / j.biocel.2014.04.020
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кристенсен, С., Пурслоу, П. П. (2016). Роль матриксных металлопротеиназ в развитии мышечной и жировой ткани и качестве мяса: обзор. Meat Sci. 119, 138–146. DOI: 10.1016 / j.meatsci.2016.04.025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коркоран, М. Л., Хьюитт, Р. Э., Кляйнер, Д.Э. младший и Стетлер-Стивенсон У. Г. (1996). ММП-2: экспрессия, активация и ингибирование. Ферментный белок 49, 7–19. DOI: 10.1159 / 000468613
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корнелисон, Д. Д., Филла, М. С., Стэнли, Х. М., Рапрэгер, А. С., и Олвин, Б. Б. (2001). Синдекан-3 и синдекан-4 специфически маркируют сателлитные клетки скелетных мышц и участвуют в поддержании сателлитных клеток и регенерации мышц. Dev. Биол. 239, 79–94.DOI: 10.1006 / dbio.2001.0416
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Косгроув, Б. Д., Гилберт, П. М., Порпилья, Э., Муркиоти, Ф., Ли, С. П., Корбел, С. Ю. и др. (2014). Омоложение популяции мышечных стволовых клеток восстанавливает силу травмированных старых мышц. Нац. Med. 20, 255–264. DOI: 10,1038 / нм 3464
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Crameri, R.M., Aagaard, P., Qvortrup, K., Langberg, H., Олесен, Дж., И Кьяер, М. (2007). Повреждение миофибрилл в скелетных мышцах человека: эффекты электростимуляции по сравнению с произвольным сокращением : равномерная по сравнению с гетерогенной активацией мышечных волокон. J. Physiol. 583, 365–380. DOI: 10.1113 / jphysiol.2007.128827
CrossRef Полный текст | Google Scholar
Крамери, Р. М., Лангберг, Х., Тейснер, Б., Магнуссон, П., Шредер, Х. Д., Олесен, Дж. Л. и др. (2004). Усиление обработки проколлагена в скелетных мышцах после однократной эксцентрической нагрузки у людей. Matrix Biol. 23, 259–264. DOI: 10.1016 / j.matbio.2004.05.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цуй, Н., Ху, М., и Халил, Р. А. (2017). Биохимические и биологические признаки матричных металлопротеиназ. Прог. Мол. Биол. Пер. Sci. 147, 1–73. DOI: 10.1016 / bs.pmbts.2017.02.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
де Соуза Нето, И. В., Дуриган, Дж. Л. К., Гуццони, В., Тибана, Р.A., Prestes, J., de Araujo, H. S. S. и др. (2018). Влияние тренировок с отягощениями на активность матричной металлопротеиназы в скелетных мышцах и кровообращение при старении. Фронт. Physiol. 9: 190. DOI: 10.3389 / fphys.2018.00190
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дитрих, А. В., Боттер, А., Виейра, Т. М., Пеолссон, А., Петцке, Ф., Дэйви, П. и др. (2017). Пространственная вариация и несоответствие между оценками начала мышечной активации по ЭМГ и УЗИ. Sci. Отчет 7: 42011. DOI: 10.1038 / srep42011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Dietrich, S., Abou-Rebyeh, F., Brohmann, H., Bladt, F., Sonnenberg-Riethmacher, E., Yamaai, T., et al. (1999). Роль SF / HGF и c-Met в развитии скелетных мышц. Dev. Camb. Англ. 126, 1621–1629.
PubMed Аннотация | Google Scholar
Duance, V.C., Restall, D.J., Beard, H., Bourne, F.J, and Bailey, A.J. (1977). Расположение трех типов коллагена в скелетных мышцах. FEBS Lett. 79, 248–252. DOI: 10.1016 / 0014-5793 (77) 80797-7
CrossRef Полный текст | Google Scholar
Эклунд, Л., Пиухола, Дж., Комулайнен, Дж., Сормунен, Р., Онгваррасопоне, К., Фасслер, Р., и др. (2001). Недостаток коллагена типа XV вызывает у мышей скелетную миопатию и сердечно-сосудистые дефекты. Proc. Natl. Акад. Sci. США 98, 1194–1199. DOI: 10.1073 / pnas.031444798
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энглер, А.Дж., Гриффин, М. А., Сен, С., Беннеман, К. Г., Суини, Х. Л. и Дишер, Д. Э. (2004). Миотрубки оптимально дифференцируются на субстратах с тканевой жесткостью: патологические последствия для мягких или жестких микроокружений. J. Cell Biol. 166, 877–887. DOI: 10.1083 / jcb.200405004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Этьен, Дж., Лю, К., Скиннер, К. М., Конбой, М. Дж., И Конбой, И. М. (2020). Скелетные мышцы как экспериментальная модель выбора для изучения старения и омоложения тканей. Скелет. Мышца 10: 4. DOI: 10.1186 / s13395-020-0222-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фишер, Дж., Стил, Дж., Брюс-Лоу, С., и Смит, Д. (2011). Рекомендации по тренировкам с отягощениями, основанные на фактах. Med. Спорт. 15, 147–162. DOI: 10.2478 / v10036-011-0025-x
CrossRef Полный текст | Google Scholar
Фолланд, Дж. П., и Уильямс, А. Г. (2007). Адаптация к силовым тренировкам: морфологический и неврологический вклад в увеличение силы. Sports Med. 37, 145–168. DOI: 10.2165 / 00007256-200737020-00004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фрай, К. С., Кирби, Т. Дж., Космак, К., Маккарти, Дж. Дж., И Петерсон, К. А. (2017). Миогенные клетки-предшественники контролируют продукцию внеклеточного матрикса фибробластами во время гипертрофии скелетных мышц. Стволовые клетки клеток 20, 56–69. DOI: 10.1016 / j.stem.2016.09.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гарбер, К.E., Blissmer, B., Deschenes, M. R., Franklin, B. A., Lamonte, M. J., Lee, I.-M., et al. (2011). Количество и качество упражнений для развития и поддержания кардиореспираторной, скелетно-мышечной и нейромоторной формы у практически здоровых взрослых: руководство по назначению упражнений. Med. Sci. Спортивные упражнения. 43, 1334–1359. DOI: 10.1249 / MSS.0b013e318213fefb
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гилберт, П. М., Хэвенстрайт, К. Л., Магнуссон, К.Э. Г., Сакко А., Леонарди Н. А., Крафт П. и др. (2010). Эластичность субстрата регулирует самообновление стволовых клеток скелетных мышц в культуре. Наука 329, 1078–1081. DOI: 10.1126 / science.11
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиллис, А. Р., Чепмен, М. А., Бушонг, Э. А., Диринк, Т. Дж., Эллисман, М. Х., и Либер, Р. Л. (2017). Трехмерная реконструкция с высоким разрешением внеклеточного матрикса фиброзных скелетных мышц: организация внеклеточного матрикса фиброзных мышц. J. Physiol. 595, 1159–1171. DOI: 10.1113 / JP273376
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Госселин, Л. Е., Адамс, К., Коттер, Т. А., Маккормик, Р. Дж., И Томас, Д. П. (1998). Влияние физических упражнений на пассивную жесткость опорно-двигательных скелетных мышц: роль внеклеточного матрикса. J. Appl. Physiol. 85, 1011–1016. DOI: 10.1152 / jappl.1998.85.3.1011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грефте, С., Vullinghs, S., Kuijpers-Jagtman, A.M., Torensma, R., and Von den Hoff, J. W. (2012). Матригель, но не коллаген I, поддерживает способность к дифференцировке мышечных клеток in vitro . Biomed. Матер. 7: 055004. DOI: 10.1088 / 1748-6041 / 7/5/055004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Grounds, M. D. (1998). Возрастные изменения реакции клеток скелетных мышц на упражнения и регенерацию. Ann. Акад.Sci. 854, 78–91. DOI: 10.1111 / j.1749-6632.1998.tb09894.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Основания, М. Д., Сорокин, Л., Уайт, Дж. (2005). Сила на границе межклеточного матрикса и мышцы. Сканд. J. Med. Sci. Спорт 15, 381–391. DOI: 10.1111 / j.1600-0838.2005.00467.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гийон, Э., Брето, С., Руджеро, Ф. (2016). Предшественники медленных мышц откладывают отпечаток пальца матрицы XV коллагена, чтобы направлять навигацию по моторным аксонам. J. Neurosci. 36, 2663–2676. DOI: 10.1523 / JNEUROSCI.2847-15.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гутьеррес, Дж., И Брандан, Э. (2010). Новый механизм связывания фактора роста фибробластов 2 с помощью глипикана в липидных рафтах, позволяющий дифференцировать скелетные мышцы. Мол. Клетка. Биол. 30, 1634–1649. DOI: 10.1128 / mcb.01164-09
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуццони, В., Рибейро, М.Б. Т., Лопес, Г. Н., де Касия Маркети, Р., де Андраде, Р. В., Селистр-де-Араужо, Х. С. и др. (2018). Влияние тренировки с отягощениями на адаптацию внеклеточного матрикса в скелетных мышцах старых крыс. Фронт. Physiol. 9: 374. DOI: 10.3389 / fphys.2018.00374
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Халпер, Дж., И Кьяер, М. (2014). «Основные компоненты соединительной ткани и внеклеточного матрикса: эластин, фибриллин, фибулины, фибриноген, фибронектин, ламинин, тенасцины и тромбоспондины», в Progress in Heritable Soft Connective Tissue Diseases , ed.Дж. Халпер, (Дордрехт: Спрингер), 31–47. DOI: 10.1007 / 978-94-007-7893-1_3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Härönen, H., Zainul, Z., Tu, H., Naumenko, N., Sormunen, R., Miinalainen, I., et al. (2017). Коллаген XIII обеспечивает пре- и постсинаптическую целостность нервно-мышечного синапса. Гум. Мол. Genet. 26, 2076–2090. DOI: 10.1093 / hmg / ddx101
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хассон П., ДеЛорье А., Беннетт, М., Григорьева, Э., Найче, Л. А., Папайоанну, В. Э. и др. (2010). Tbx4 и tbx5, действующие в соединительной ткани, необходимы для формирования рисунка мышц конечностей и сухожилий. Dev. Cell 18, 148–156. DOI: 10.1016 / j.devcel.2009.11.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хаус, Дж. М., Карритерс, Дж. А., Кэрролл, К. К., Теш, П. А., и Трапп, Т. А. (2007a). Содержание белка сократительной и соединительной ткани в скелетных мышцах человека: эффекты 35- и 90-дневного моделирования микрогравитации и контрмеры при физической нагрузке. Am. J. Physiol. Regul. Интегр. Комп. Physiol. 293, R1722 – R1727. DOI: 10.1152 / ajpregu.00292.2007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хаус, Дж. М., Карритерс, Дж. А., Траппе, С. У. и Траппе, Т. А. (2007b). Коллаген, сшивание и конечные продукты гликирования в стареющих скелетных мышцах человека. J. Appl. Physiol. 103, 2068–2076. DOI: 10.1152 / japplphysiol.00670.2007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хейккинен, А., Härönen, H., Norman, O., and Pihlajaniemi, T. (2019). Коллаген XIII и другие компоненты ECM в сборке и заболевании нервно-мышечного соединения. Анат. Рек. doi: 10.1002 / ar.24092 [Epub перед печатью].
CrossRef Полный текст | PubMed Аннотация | Google Scholar
Heinemeier, K. M., Olesen, J. L., Haddad, F., Langberg, H., Kjaer, M., Baldwin, K. M., et al. (2007). Экспрессия коллагена и связанных с ним факторов роста в сухожилиях и скелетных мышцах крысы в ответ на определенные типы сокращения: экспрессия коллагена и TGF-β-1 в тренированных сухожилиях и мышцах. J. Physiol. 582, 1303–1316. DOI: 10.1113 / jphysiol.2007.127639
CrossRef Полный текст | Google Scholar
Heljasvaara, R., Aikio, M., Ruotsalainen, H., and Pihlajaniemi, T. (2017). Коллаген XVIII в гомеостазе и нарушении регуляции тканей — уроки, извлеченные из модельных организмов и пациентов-людей. Matrix Biol. 57–58, 55–75. DOI: 10.1016 / j.matbio.2016.10.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хидзиката, Т., Вакисака Х. и Ниида С. (1993). Функциональная комбинация сужающихся профилей и перекрывающихся расположений в нерасширяющихся волокнах скелетных мышц, оканчивающихся внутрипучкообразно. Анат. Рек. 236, 602–610. DOI: 10.1002 / ar.10
403
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хиндл, А.Г., Хорнинг, М., Меллиш, Дж .-А. Э. и Лоулер Дж. М. (2009). Погружение в старость: мускульное старение у крупнотелого долгоживущего млекопитающего, тюленя Уэдделла ( Leptonychotes weddellii ). J. Exp. Биол. 212, 790–796. DOI: 10.1242 / jeb.025387
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холм Л., Рахбек С. К., Фаруп Дж., Вендельбо М. Х. и Виссинг К. (2017). Режим сокращения и потребление сывороточного протеина влияют на скорость синтеза внутримышечной соединительной ткани: краткие отчеты. Мышечный нерв 55, 128–130. DOI: 10.1002 / mus.25398
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холм, Л., ван Холл, Г., Роуз, А. Дж., Миллер, Б. Ф., Дёссинг, С., Рихтер, Э. А. и др. (2010). Интенсивность сокращения и питание по-разному влияют на скорость синтеза коллагена и миофибриллярного белка в скелетных мышцах человека. Am. J. Physiol. Эндокринол. Метаб. 298, E257 – E269. DOI: 10.1152 / ajpendo.00609.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хуанг, Ю., Фан, Ю., Саланова, М., Янг, X., Сан, Л., и Блоттнер, Д. (2018). Влияние подошвенной вибрации на кости и глубокую фасцию в модели неиспользования разгрузки задней конечности крысы. Фронт. Physiol. 9: 616. DOI: 10.3389 / fphys.2018.00616
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хьюз, Д. К., Маркотт, Г. Р., Маршалл, А. Г., Уэст, Д. В. Д., Баер, Л. М., Уоллес, М. А., и др. (2016). Возрастные различия дистрофина: влияние на белки передачи силы, целостность мембран и стабильность нервно-мышечных соединений. J. Gerontol. A. Biol. Sci. Med. Sci. 72, 640–648. DOI: 10.1093 / gerona / glw109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хуцзин, П.А. (1999). Мышцы как композит, армированный коллагеновыми волокнами: обзор передачи силы в мышце и всей конечности. J. Biomech. 32, 329–345. DOI: 10.1016 / s0021-9290 (98) 00186-9
CrossRef Полный текст | Google Scholar
Хьюцзин П.А., Баан Г.С. и Ребел Г.Т. (1998). Немышухожильная передача силы в длинном разгибателе пальцев большого пальца крысы. J. Exp. Биол. 201, 683–691.
Google Scholar
Хилдал, Р. Д., Нельсон, Б., Xin, L., Welling, T., Groscost, L., Hubal, M.J., et al. (2015). Ремоделирование внеклеточного матрикса и его вклад в защитную адаптацию после удлинения сокращений в мышцах человека. FASEB J. 29, 2894–2904. DOI: 10.1096 / fj.14-266668
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ивата, Дж., Судзуки, А., Пеликан, Р. К., Хо, Т.-В., и Чай, Ю. (2013). Передача сигналов неканонического трансформирующего фактора роста β (TGFβ) в клетках краниального нервного гребня вызывает дефекты развития мышц языка. J. Biol. Chem. 288, 29760–29770. DOI: 10.1074 / jbc.M113.493551
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Якобсен, Дж. Р., Макки, А. Л., Кнудсен, А. Б., Кох, М., Кьяер, М., и Крогсгаард, М. Р. (2017). Состав и адаптация мышечно-сухожильных суставов человека и соседних мышечных волокон к тяжелым тренировкам с отягощениями. Сканд. J. Med. Sci. Спорт 27, 1547–1559. DOI: 10.1111 / sms.12794
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Яначек, Я., Цебашек, В., Кубинова, Л., Рибарич, С., и Эрзен, И. (2009). Трехмерная визуализация и измерение капилляров, снабжающих метаболически различные типы волокон в длинном разгибателе пальцев большого пальца крысы во время денервации и реиннервации. J. Histochem. Cytochem. 57, 437–447. DOI: 10.1369 / jhc.2008.953018
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ярвинен, Т.А. Х., Йожа, Л., Каннус, П., Ярвинен, Т. Л. Н., и Ярвинен, М. (2002). Организация и распределение внутримышечной соединительной ткани в нормальных и неподвижных скелетных мышцах.Иммуногистохимическое, поляризационное и сканирующее электронно-микроскопические исследования. J. Muscle Res. Cell Motil. 23, 245–254. DOI: 10.1023 / A: 10208336
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Канг, Л., Лантье, Л., Кеннеди, А., Боннер, Дж. С., Мэйс, В. Х., Брейси, Д. П. и др. (2013). Гиалуронан накапливается при кормлении с высоким содержанием жиров и способствует развитию инсулинорезистентности. Диабет 62, 1888–1896. DOI: 10.2337 / db12-1502
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Канг, Л., Мэйс, У. Х., Джеймс, Ф. Д., Брейси, Д. П., и Вассерман, Д. Х. (2014). Матричная металлопротеиназа 9 противодействует индуцированной диетой мышечной инсулинорезистентности у мышей. Диабетология 57, 603–613. DOI: 10.1007 / s00125-013-3128-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кардон, Г., Харф, Б. Д. и Табин, К. Дж. (2003). Tcf4-позитивная мезодермальная популяция обеспечивает предпаттерн для формирования мышечного паттерна конечностей позвоночных. Dev. Cell 5, 937–944. DOI: 10.1016 / с 1534-5807 (03) 00360-5
CrossRef Полный текст | Google Scholar
Карпакка, Й., Вяэнянен, К., Орава, С., и Такала, Т. Э. (1990). Эффекты предварительной иммобилизации и иммобилизации на синтез коллагена в скелетных мышцах крыс. Внутр. J. Sports Med. 11, 484–488. DOI: 10.1055 / с-2007-1024842
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Карпакка, Дж., Виртанен, П., Ваананен, К., Орава, С., и Такала, Т. Е. (1991).Синтез коллагена в скелетных мышцах крысы во время иммобилизации и ремобилизации. J. Appl. Physiol. 70, 1775–1780. DOI: 10.1152 / jappl.1991.70.4.1775
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Khaleduzzaman, M., Sumiyoshi, H., Ueki, Y., Inoguchi, K., Ninomiya, Y., and Yoshioka, H. (1997). Структура гена коллагена человека типа XIX (COL19A1), которая предполагает, что он произошел от гена-предка семейства FACIT. Геномика 45, 304–312.DOI: 10.1006 / geno.1997.4921
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ким Дж., Ван З., Хеймсфилд С. Б., Баумгартнер Р. Н. и Галлахер Д. (2002). Общая масса скелетных мышц: оценка с помощью нового метода двухэнергетической рентгеновской абсорбциометрии. Am. J. Clin. Nutr. 76, 378–383. DOI: 10.1093 / ajcn / 76.2.378
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кнудсен, А.Б., Ларсен, М., Макки, А.Л., Хьорт, М., Hansen, K. K., Qvortrup, K., et al. (2015). Человеческое мышечно-сухожильное соединение: ультраструктурное исследование и трехмерный анализ. Сканд. J. Med. Sci. Sports 25, e116 – e123. DOI: 10.1111 / sms.12221
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Koch, M., Schulze, J., Hansen, U., Ashwodt, T., Keene, D.R., Brunken, W.J., et al. (2004). Новый маркер тканевых соединений, коллаген XXII. J. Biol. Chem. 279, 22514–22521. DOI: 10.1074 / jbc.M400536200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кованен, В., Суоминен, Х., Ристели, Дж., И Ристели, Л. (1988). Коллаген IV типа и ламинин в медленных и быстрых скелетных мышцах крыс — Влияние возраста и тренировок на выносливость на протяжении всей жизни. Сб. Relat. Res. 8, 145–153. DOI: 10,1016 / s0174-173x (88) 80026-8
CrossRef Полный текст | Google Scholar
Куо, Х. Дж., Маслен, К. Л., Кин, Д. Р., и Гланвилл, Р. У. (1997). Коллаген VI типа закрепляет базальные мембраны эндотелия, взаимодействуя с коллагеном IV типа. J. Biol. Chem. 272, 26522–26529. DOI: 10.1074 / jbc.272.42.26522
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Леонг, Д. П., Тео, К. К., Рангараджан, С., Лопес-Харамилло, П., Авесум, А., Орландини, А., и др. (2015). Прогностическое значение силы хвата: результаты проспективного исследования городской и сельской эпидемиологии (PURE). Lancet Lond. Англ. 386, 266–273. DOI: 10,1016 / s0140-6736 (14) 62000-6
CrossRef Полный текст | Google Scholar
Ли, Дж.Дж., Виттерт, Г. А., Винсент, А., Атлантис, Э., Ши, З., Эпплтон, С. Л. и др. (2016). Сила мышечного захвата позволяет прогнозировать развитие диабета 2 типа: популяционное когортное исследование. Метаболизм 65, 883–892. DOI: 10.1016 / j.metabol.2016.03.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Либер, Р. Л., и Уорд, С. Р. (2013). Клеточные механизмы тканевого фиброза. 4. Структурные и функциональные последствия фиброза скелетных мышц. Am. J. Physiol.Cell Physiol. 305, C241 – C252. DOI: 10.1152 / ajpcell.00173.2013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лопес-Отин, К., Бласко, М.А., Партридж, Л., Серрано, М., и Кремер, Г. (2013). Признаки старения. Ячейка 153, 1194–1217. DOI: 10.1016 / j.cell.2013.05.039
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Maaß, T., Bayley, C.P., Mörgelin, M., Lettmann, S., Bonaldo, P., Paulsson, M., et al. (2016).Неоднородность микрофибрилл коллагена VI: структурный анализ неколлагеновых участков. J. Biol. Chem. 291, 5247–5258. DOI: 10.1074 / jbc.M115.705160
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mackey, A. L., Brandstetter, S., Schjerling, P., Bojsen-Moller, J., Qvortrup, K., Pedersen, M. M., et al. (2011). Последовательный ответ мертвых клеток внеклеточного матрикса и фиброзных регуляторов после повреждения мышц участвует в защите скелетных мышц человека от будущих повреждений. FASEB J. 25, 1943–1959. DOI: 10.1096 / fj.10-176487
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Mackey, A. L., Donnelly, A. E., Turpeenniemi-Hujanen, T., and Roper, H.P. (2004). Содержание коллагена в скелетных мышцах у людей после эксцентрических сокращений большой силы. J. Appl. Physiol. 97, 197–203. DOI: 10.1152 / japplphysiol.01174.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Махновский П.А., Згода В.Г., Боков Р.О., Шагимарданова Е.И., Газизова Г.Р., Гусев О.А. и др. (2020). Регуляция белков в скелетных мышцах человека: роль транскрипции. Sci. Отчет 10: 3514.
Google Scholar
Мартин, Г. Р., и Тимпл, Р. (1987). Ламинин и другие компоненты базальной мембраны. Annu. Rev. Cell Biol. 3, 57–85. DOI: 10.1146 / annurev.cb.03.110187.000421
CrossRef Полный текст | Google Scholar
Макки, Т. Дж., Перлман, Г., Моррис, М., Комарова, С. В. (2019). Состав внеклеточного матрикса соединительной ткани: систематический обзор и метаанализ. Sci. Отчет 9: 10542.
Google Scholar
Мендиас, К. Л., Шварц, А. Дж., Грекин, Дж. А., Гумусио, Дж. П., и Сугг, К. Б. (2017). Изменения сократимости мышечных волокон и продукции внеклеточного матрикса при гипертрофии скелетных мышц. J. Appl. Physiol. 122, 571–579. DOI: 10.1152 / japplphysiol.00719.2016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монти, Р.Дж., Рой, Р. Р., Ходжсон, Дж. А., и Реджи Эджертон, В. (1999). Передача сил в скелетных мышцах млекопитающих. J. Biomech. 32, 371–380. DOI: 10.1016 / s0021-9290 (98) 00189-4
CrossRef Полный текст | Google Scholar
Мур, Д. Р., Филлипс, С. М., Бабрадж, Дж. А., Смит, К., и Ренни, М. Дж. (2005). Синтез миофибриллярных и коллагеновых белков в скелетных мышцах человека у молодых мужчин после максимального сокращения и удлинения сокращений. Am. J. Physiol.Эндокринол. Метаб. 288, E1153 – E1159. DOI: 10.1152 / ajpendo.00387.2004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моралес, М. Г., Кабелло-Верругио, К., Сантандер, К., Кабрера, Д., Гольдшмединг, Р., и Брандан, Э. (2011). Сверхэкспрессия CTGF / CCN-2 может напрямую вызывать признаки дистрофии скелетных мышц. J. Pathol. 225, 490–501. DOI: 10.1002 / путь.2952
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Малви, К., Харно, Э., Кинан, А., и Олендик, К. (2005). Экспрессия комплекса дистрофин-дистрогликан скелетных мышц и комплекса синтрофин-оксид азота сильно нарушена у крыс Goto-Kakizaki с диабетом 2 типа. Eur. J. Cell Biol. 84, 867–883. DOI: 10.1016 / j.ejcb.2005.06.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Наба А., Клаузер К. Р., Динг Х., Уиттакер К. А., Карр С. А. и Хайнс Р. О. (2016). Внеклеточный матрикс: инструменты и идеи для эпохи «омиков». Matrix Biol. 49, 10–24. DOI: 10.1016 / j.matbio.2015.06.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нассари, С., Дюпре, Д., Фурнье-Тибо, К. (2017). Немиогенный вклад в развитие мышц и гомеостаз: роль соединительной ткани. Фронт. Cell Dev. Биол. 5:22. DOI: 10.3389 / fcell.2017.00022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Nordez, A., Gallot, T., Catheline, S., Гевель А., Корню К. и Хуг Ф. (2009). Электромеханическая задержка пересмотрена с использованием ультразвука с очень высокой частотой кадров. J. Appl. Physiol. 106, 1970–1975. DOI: 10.1152 / japplphysiol.00221.2009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Новицки Дж. Л., Такимото Р. и Берк А. С. (2003). Латеральная сомитная граница: дорсо-вентральные аспекты передне-задней регионализации у эмбрионов птиц. мех. Dev. 120, 227–240. DOI: 10.1016 / s0925-4773 (02) 00415-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Огасавара, Р., Наказато К., Сато К., Боппарт М. Д. и Фудзита С. (2014). Упражнения с отягощениями увеличивают экспрессию активного белка MMP и бета1-интегрина в скелетных мышцах. Physiol. Отчет 2: e12212. DOI: 10.14814 / phy2.12212
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Olguin, H.C., и Olwin, B.B. (2004). Повышающая регуляция Pax-7 ингибирует миогенез и прогрессию клеточного цикла в сателлитных клетках: потенциальный механизм самообновления. Dev. Биол. 275, 375–388.DOI: 10.1016 / j.ydbio.2004.08.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Олссон, Л., Фальк, П., Лопес, К., Кобб, Дж., И Ханкен, Дж. (2001). Клетки черепного нервного гребня вносят вклад в соединительную ткань в черепных мышцах бесхвостых амфибий, Bombina orientalis . Dev. Биол. 237, 354–367. DOI: 10.1006 / dbio.2001.0377
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Passerieux, E., Rossignol, R., Летелье Т. и Делаж Дж. (2007). Физическая непрерывность перимизиума от миофибрилл к сухожилиям: участие в передаче латеральной силы в скелетных мышцах. J. Struct. Биол. 159, 19–28. DOI: 10.1016 / j.jsb.2007.01.022
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паттисон, Дж. С., Фолк, Л. К., Мэдсен, Р. У., Чайлдс, Т. Э. и Бут, Ф. У. (2003). Транскрипционное профилирование выявляет обширное подавление экспрессии гена внеклеточного матрикса в саркопенической камбаловидной мышце крысы. Physiol. Геномика 15, 34–43. DOI: 10.1152 / Physiolgenomics.00040.2003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Паван, П. Г., Стекко, А., Стерн, Р., и Стекко, К. (2014). Болезненные связи: уплотнение или фиброз фасции. Curr. Боль Головная боль Отчет 18: 441.
Google Scholar
Пирс Р. В., Шерц П. Дж., Кэмпбелл Дж. К. и Табин К. Дж. (2007). Анализ клеточного происхождения зачатка куриной конечности. Dev. Биол. 310, 388–400. DOI: 10.1016 / j.ydbio.2007.08.002
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Питер А. К., Ченг Х., Росс Р. С., Ноултон К. У. и Чен Дж. (2011). Костамер соединяет саркомеры с сарколеммой поперечно-полосатой мышцы. Прог. Педиатр. Кардиол. 31, 83–88. DOI: 10.1016 / j.ppedcard.2011.02.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перслоу П. П. (2002). Структура и функциональное значение изменений соединительной ткани в мышце. Сост. Биохим. Physiol. Мол. Интегр. Physiol. 133, 947–966. DOI: 10.1016 / s1095-6433 (02) 00141-1
CrossRef Полный текст | Google Scholar
Рамасвами, К. С., Палмер, М. Л., ван дер Мейлен, Дж. Х., Рену, А., Костроминова, Т. Ю., Мишель, Д. Е. и др. (2011). Боковая передача силы нарушена в скелетных мышцах дистрофических мышей и очень старых крыс: латеральная передача силы в скелетных мышцах мышей и крыс. J. Physiol. 589, 1195–1208.DOI: 10.1113 / jphysiol.2010.201921
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райагири С.С., Ранальди Д., Рэйвен А., Мохамад Азхар Н. И. Ф., Лефевр О., Заммит П. С. и др. (2018). Ремоделирование базальной пластинки в нише стволовых клеток скелетных мышц опосредует самообновление стволовых клеток. Нац. Commun. 9: 1075.
Google Scholar
Риз, С. П., Андервуд, К. Дж., И Вайс, Дж. А. (2013). Влияние протеогликана декорина на фибриллогенез, ультраструктуру и механику коллагеновых гелей типа I. Matrix Biol. 32, 414–423. DOI: 10.1016 / j.matbio.2013.04.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Реннинг, С. Б., Педерсен, М. Э., Андерсен, П. В., и Холлунг, К. (2013). Комбинация гликозаминогликанов и фиброзных белков улучшает пролиферацию клеток и раннюю дифференциацию первичных клеток скелетных мышц крупного рогатого скота. Дифференциация 86, 13–22. DOI: 10.1016 / j.diff.2013.06.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роуленд, Л.А., Бал, Н. К., Периасами, М. (2015). Роль термогенных механизмов скелетных мышц в эндотермии позвоночных. Biol. Преподобный Камб. Филос. Soc. 90, 1279–1297. DOI: 10.1111 / brv.12157
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рубенштейн, А. Б., Смит, Г. Р., Рауэ, У., Бег, Г., Минчев, К., Руф-Замойски, Ф. и др. (2020). Одноклеточные профили транскрипции в скелетных мышцах человека. Sci. Отчет 10: 229.
Google Scholar
Сабателли, П., Гуаланди, Ф., Гара, С. К., Грумати, П., Зампарелли, А., Мартони, Э. и др. (2012). Экспрессия α5 и α6 цепей коллагена VI в мышцах человека и при мышечном фиброзе, связанном с мышечной дистрофией Дюшенна. Matrix Biol. 31, 187–196. DOI: 10.1016 / j.matbio.2011.12.003
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Санес, Дж. Р. (1982). Ламинин, фибронектин и коллаген в синаптических и внесинаптических частях базальной мембраны мышечных волокон. Дж.Cell Biol. 93, 442–451. DOI: 10.1083 / jcb.93.2.442
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Саволайнен, Дж., Вяэнянен, К., Пуранен, Дж., Такала, Т. Э., Комулайнен, Дж., И Вихко, В. (1988). Синтез коллагена и протеолитическая активность в скелетных мышцах крыс: эффект гипсовой иммобилизации в удлиненных и укороченных положениях. Arch. Phys. Med. Rehabil. 69, 964–969.
PubMed Аннотация | Google Scholar
Шенфельд, Б.Дж. (2010). Механизмы гипертрофии мышц и их применение в тренировках с отягощениями. J. Strength Cond. Res. 24, 2857–2872. DOI: 10.1519 / JSC.0b013e3181e840f3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шенрок Б., Зандер В., Дерн С., Лимпер У., Малдер Э., Веракситш А. и др. (2018). Постельный режим, противодействие физическим упражнениям и восстанавливающее воздействие на систему мышечного тонуса человека в состоянии покоя. Фронт. Physiol. 9: 810. DOI: 10.3389 / fphys.2018.00810
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Schönherr, E., Witsch-Prehm, P., Harrach, B., Robenek, H., Rauterberg, J., and Kresse, H. (1995). Взаимодействие бигликана с коллагеном I типа. J. Biol. Chem. 270, 2776–2783. DOI: 10.1074 / jbc.270.6.2776
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шварц Р. И. (2015). Коллаген I и фибробласты: высокая экспрессия белка требует новой парадигмы посттранскрипционной регуляции с обратной связью. Biochem. Биофиз. Rep. 3, 38–44. DOI: 10.1016 / j.bbrep.2015.07.007
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сефтон, Э. М., и Кардон, Г. (2019). Связующее развитие мышц, врожденные дефекты и эволюция: важная роль соединительной ткани мышц. Curr. Вершина. Dev. Биол. 132, 137–176. DOI: 10.1016 / bs.ctdb.2018.12.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шарафи Б. и Блемкер С.С. (2011). Математическая модель передачи силы от оканчивающихся внутри ячеек мышечных волокон. J. Biomech. 44, 2031–2039. DOI: 10.1016 / j.jbiomech.2011.04.038
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Соренсен, Дж. Р., Скоузен, К., Холланд, А., Уильямс, К., и Хилдал, Р. Д. (2018). Острый внеклеточный матрикс, воспалительная реакция и реакция MAPK на удлинение сокращений в скелетных мышцах пожилого человека. Exp. Геронтол. 106, 28–38.DOI: 10.1016 / j.exger.2018.02.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Street, S. F. (1983). Боковая передача напряжения в миофибриллах лягушки: миофибриллярная сеть и поперечные цитоскелетные связи являются возможными передатчиками. J. Cell. Physiol. 114, 346–364. DOI: 10.1002 / jcp.1041140314
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сумиёси, Х., Лауб, Ф., Йошиока, Х., и Рамирес, Ф. (2001).Эмбриональная экспрессия коллагена типа XIX преходяща и ограничена мышечными клетками. Dev. Дин. 220, 155–162. DOI: 10.1002 / 1097-0177 (2000) 9999: 9999 <:: aid-dvdy1099> 3.0.co; 2-w
CrossRef Полный текст | Google Scholar
Суоминен, Х., Хейккинен, Э. (1975). Активность ферментов в мышечной и соединительной ткани M. Vastus lateralis у мужчин от 33 до 70 лет, которые обычно тренируются и ведут малоподвижный образ жизни. Eur. J. Appl. Physiol. 34, 249–254. DOI: 10.1007 / bf00999938
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Суоминен, Х., Хейккинен, Э., и Паркатти, Т. (1977). Влияние восьминедельной физической тренировки на мышцы и соединительную ткань M. Vastus lateralis у 69-летних мужчин и женщин. J. Gerontol. 32, 33–37. DOI: 10.1093 / geronj / 32.1.33
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Swartz, M. E., Eberhart, J., Pasquale, E. B., and Krull, C.Э. (2001). Взаимодействия EphA4 / эфрин-A5 при миграции клеток-предшественников мышц в передних конечностях птиц. Dev. Camb. Англ. 128, 4669–4680.
PubMed Аннотация | Google Scholar
Такаги Р., Огасавара Р., Цутаки А., Наказато К. и Исии Н. (2016). Региональная адаптация коллагена в скелетных мышцах к повторяющимся сериям напряженных эксцентрических упражнений. Pflugers Arch. 468, 1565–1572. DOI: 10.1007 / s00424-016-1860-3
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такала, Т.Э. и Виртанен П. (2000). Биохимический состав внеклеточного матрикса мышц: эффект нагрузки. Сканд. J. Med. Sci. Спорт 10, 321–325. DOI: 10.1034 / j.1600-0838.2000.010006321.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Takala, T. E. S., Myllylä, R., Salminen, A., Anttinen, H., and Vihko, V. (1983). Повышенная активность пролил-4-гидроксилазы и галактозилгидроксилизилглюкозилтрансферазы, ферментов биосинтеза коллагена, в скелетных мышцах мышей, тренированных на выносливость. Pflügers Arch. 399, 271–274. DOI: 10.1007 / BF00652751
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Там, К. С., Чаудхури, Р., Хатчисон, А. Т., Самоча-Бонет, Д., и Хайльбронн, Л. К. (2017). Ремоделирование внеклеточного матрикса скелетных мышц после кратковременного переедания у здоровых людей. Метаболизм 67, 26–30. DOI: 10.1016 / j.metabol.2016.10.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Там, К.С., Ковингтон, Дж. Д., Баджпей, С., Чукалова, Ю., Бурк, Д., Йоханнсен, Д. Л. и др. (2014). Увеличение веса показывает резкое увеличение ремоделирования внеклеточного матрикса скелетных мышц. J. Clin. Эндокринол. Метаб. 99, 1749–1757. DOI: 10.1210 / jc.2013-4381
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Tam, C. S., Power, J. E., Markovic, T. P., Yee, C., Morsch, M., McLennan, S. V., et al. (2015). Влияние питания с высоким содержанием жиров на физическую функцию и внеклеточный матрикс скелетных мышц. Nutr. Диабет 5: e187. DOI: 10.1038 / nutd.2015.39
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Thorsteinsdóttir, S., Deries, M., Cachaço, A. S., and Bajanca, F. (2011). Размер внеклеточного матрикса развития скелетных мышц. Dev. Биол. 354, 191–207. DOI: 10.1016 / j.ydbio.2011.03.015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тирни, М. Т., Громова, А., Сесилло, Ф. Б., Сала, Д., Спенле, К., Orend, G., et al. (2016). Автономное ремоделирование внеклеточного матрикса контролирует прогрессивную адаптацию регенеративной способности мышечных стволовых клеток во время развития. Cell Rep. 14, 1940–1952. DOI: 10.1016 / j.celrep.2016.01.072
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уэдзуми А., Фукада С., Ямамото Н., Такеда С. и Цучида К. (2010). Мезенхимальные предшественники, отличные от сателлитных клеток, способствуют образованию эктопических жировых клеток в скелетных мышцах. Нац. Cell Biol. 12, 143–152. DOI: 10.1038 / ncb2014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Uhlen, M., Oksvold, P., Fagerberg, L., Lundberg, E., Jonasson, K., Forsberg, M., et al. (2010). К атласу белков человека, основанному на знаниях. Нац. Biotechnol. 28, 1248–1250. DOI: 10.1038 / nbt1210-1248
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Урчиуоло, А., Куарта, М., Морбидони, В., Гаттаццо, Ф., Молон С., Грумати П. и др. (2013). Коллаген VI регулирует самообновление сателлитных клеток и регенерацию мышц. Нац. Commun. 4: 1964. DOI: 10.1038 / ncomms2964
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Vallecillo-García, P., Orgeur, M., Vom Hofe-Schneider, S., Stumm, J., Kappert, V., Ibrahim, D. M., et al. (2017). Odd skipped-related 1 идентифицирует популяцию эмбриональных фибро-адипогенных предшественников, регулирующих миогенез во время развития конечностей. Нац. Commun. 8: 1218.
Google Scholar
Вирджилио, К. М., Мартин, К. С., Пирс, С. М., и Блемкер, С. С. (2015). Многомасштабные модели скелетных мышц показывают комплексное влияние мышечной дистрофии на механику тканей и восприимчивость к повреждениям. Интерфейс Focus 5: 20140080. DOI: 10.1098 / rsfs.2014.0080
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Visse, R., and Nagase, H. (2003). Матричные металлопротеиназы и тканевые ингибиторы металлопротеиназ: структура, функция и биохимия. Circ. Res. 92, 827–839. DOI: 10.1161 / 01.RES.0000070112.80711.3D
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, D., Eraslan, B., Wieland, T., Hallström, B., Hopf, T., Zolg, D. P., et al. (2019). Атлас обилия протеома и транскриптомов 29 здоровых тканей человека. Мол. Syst. Биол. 15: e8503. DOI: 10.15252 / msb.20188503
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Весснер, Б., Либенштайнер, М., Nachbauer, W., and Csapo, R. (2019). Возрастной ответ внеклеточного матрикса скелетных мышц на упражнения с отягощениями: пилотное исследование. Eur. J. Sport Sci. 19, 354–364. DOI: 10.1080 / 17461391.2018.1526974
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уайт, Дж. П., Риси, Дж. М., Вашингтон, Т. А., Сато, С., Ле, М. Е., Дэвис, Дж. М. и др. (2009). Вызванное перегрузкой ремоделирование внеклеточного матрикса скелетных мышц и рост миофибрилл у мышей, лишенных IL-6. Acta Physiol. 197, 321–332. DOI: 10.1111 / j.1748-1716.2009.02029.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вильшут, К. Дж., Хаагсман, Х. П., и Ролен, Б. А. Дж. (2010). Компоненты внеклеточного матрикса определяют поведение стволовых клеток мускулов свиньи. Exp. Cell Res. 316, 341–352. DOI: 10.1016 / j.yexcr.2009.10.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wood, L. K., Kayupov, E., Gumucio, J. P., Мендиас, К. Л., Клафлин, Д. Р., Брукс, С. В. (2014). Внутренняя жесткость внеклеточного матрикса в скелетных мышцах мышей увеличивается с возрастом. J. Appl. Physiol. 117, 363–369. DOI: 10.1152 / japplphysiol.00256.2014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Yucesoy, C.A., Maas, H., Koopman, B.H.F.J.M., Grootenboer, H.J., и Huijing, P.A. (2006). Механизмы, влияющие на положение мышц на проксимо-дистальные различия мышечной силы при передаче внемышечной миофасциальной силы. Med. Англ. Phys. 28, 214–226. DOI: 10.1016 / j.medengphy.2005.06.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Заммит П. С. (2017). Функция миогенных регуляторных факторов Myf5, MyoD, Myogenin и MRF4 в скелетных мышцах, сателлитных клетках и регенеративном миогенезе. Семин. Cell Dev. Биол. 72, 19–32. DOI: 10.1016 / j.semcdb.2017.11.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан, К., и Гао, Ю. (2012). Конечно-элементный анализ механики боковой передачи силы в одиночном мышечном волокне. J. Biomech. 45, 2001–2006. DOI: 10.1016 / j.jbiomech.2012.04.026
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zhu, J., Li, Y., Shen, W., Qiao, C., Ambrosio, F., Lavasani, M., et al. (2007). Взаимосвязь между трансформирующим фактором роста-β1, миостатином и декорином: последствия для фиброза скелетных мышц. J. Biol. Chem. 282, 25852–25863.DOI: 10.1074 / jbc.M704146200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zong, H., Bastie, C.C., Xu, J., Fassler, R., Campbell, K.P., Kurland, I.J., et al. (2009). Резистентность к инсулину у мышей с дефицитом бета1-рецептора интегрина, специфичного для поперечно-полосатых мышц. J. Biol. Chem. 284, 4679–4688. DOI: 10.1074 / jbc.M807408200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цюгель, М., Маганарис, К. Н., Вильке, Дж., Юркат-Ротт, К., Klingler, W., Wearing, S.C., et al. (2018). Исследование фасциальных тканей в спортивной медицине: от молекул до адаптации тканей, травм и диагностики: консенсусное заявление. Br. J. Sports Med. 52: 1497. DOI: 10.1136 / bjsports-2018-099308
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Объяснение мышечной системы за 6 минут
Наша основная концепция здесь, на CTEskills.com, — предоставить вам необходимую информацию в ясной, краткой и точной форме.Это действительно отражено в одном из наших последних видео.
Мышечная система за 6 минут.
Мышечная система действительно очень сложна, как и все системы организма, но с помощью этого короткого видео вы должны хотя бы иметь общее представление о том, что составляет мышечную систему, ее свойствах и функциях.
Для более глубокого изучения мышечной системы ищите будущие видеоролики CTE, в которых мы рассмотрим такие темы, как 14 основных групп мышц, а также распространенные заболевания и состояния, характерные для мышечной системы.
Мышечная система — обзор
Мышечная система состоит из более 600 мышц. Хотя в этом обзорном видео мы не будем рассматривать все 600 с лишним отдельных мышц, мы будем обсуждать…
- основные функции мышечной системы,
- 5 типов мышечных движений
- и как все это вместе составляет мышечную систему.
Основное назначение мышечной системы — обеспечение движения тела.Мышцы получают способность перемещать тело через нервную систему.
Основные свойства мышечной системы
Мышечная система имеет 5 основных свойств
- Возбудимый или раздражительный: Мышцы возбудимы или раздражительны. Это означает, что они способны получать стимуляцию и реагировать на стимуляцию нервов.
- Контрактируемые: Они могут быть сокращены. После стимуляции они могут сокращаться или сокращаться.
- Extensible: Растяжимость означает, что мышца может быть растянута без повреждения путем приложения силы.
- Эластичность: Обладая эластичностью, мышца может возвращаться к своей исходной форме покоя и длине после растяжения или сокращения.
- Приспособляемость: Мышечная система приспосабливаема в том смысле, что ее можно изменять в зависимости от того, как она используется. Например, мышца увеличится или подвергнется гипертрофии при увеличении нагрузки; но с другой стороны, он может атрофироваться или исчезнуть, если его лишить работы.
Типы мышечных движений
Теперь давайте посмотрим на 5 типов мышечных движений.
- Приведение … это перемещение части тела к средней линии тела.
- Похищение … отводит часть тела от тела.
- Сгибание … Сгибание означает сгибание сустава для уменьшения угла между двумя костями или двумя частями тела.
- Разгибание … разгибание — это выпрямление и разгибание сустава для увеличения угла между двумя костями или частями тела.
- Вращение … и, наконец, вращение включает перемещение части тела вокруг оси.
3 типа мышц
Мышечная система делится на три основных типа. Каждый из этих типов можно перемещать одним из двух способов. либо добровольно, либо непроизвольно.
- Сердечная мышца — Сердечная мышца — непроизвольная мышца; это означает, что он действует без какого-либо сознательного контроля.
- Висцеральные или гладкие мышцы также считаются непроизвольными — эти мышцы находятся в органах или системах органов, таких как пищеварительная или дыхательная система.
- Третий тип мышц — Скелетная мышца . Это то, о чем мы обычно думаем, когда говорим о мышцах. Эти мышцы прикрепляются к скелету и дают скелету возможность двигаться. Скелетные мышцы классифицируются как произвольные. Это потому, что мы должны сделать сознательное усилие или принять решение, чтобы заставить их двигаться.
Опорные конструкции
Хотя мышцы, очевидно, составляют мышечную систему, все же есть некоторые поддерживающие структуры, на которые нам нужно обратить внимание, которые также являются необходимой частью системы.Сухожилия и фасции являются важными поддерживающими структурами для скелетных мышц.
Проще говоря, сухожилия — это то, что прикрепляет мышцу к кости. Фасции соединяют мышцы с другими мышцами. Сухожилия и фасции работают вместе с мышцами, создавая мышечную систему, необходимую для движения.
Нервная система
Хотя нервная система технически не является частью одной и той же системы, она неразрывно связана с мышечной системой тела.Это важно для произвольных скелетных мышц.
Если связь нервной системы со скелетными мышцами прервана, скелетные мышцы не смогут производить движения для тела. Тело сейчас парализовано. Это называется квадриплегией. Единственный орган, который напрямую не зависит от нервной системы, — это сердечная мышца.
RECAP:
В человеческом теле более 600 сотен мускулов.
Мышечная система имеет 5 основных свойств.
- Возбудимый или раздражительный
- Контракт
- Расширяемый
- Эластичность
- Адаптивность
Есть 5 типов движений мышц.
Есть 3 типа мышц; добровольное и недобровольное
- Сердечная мышца — непроизвольная
- Висцеральные или гладкие мышцы — непроизвольные
- Скелетная мышца — произвольная
Опорные конструкции
Сухожилия и фасции являются важными поддерживающими структурами для скелетных мышц.
Нервная система неразрывно связана с мышечной системой тела. Это необходимо для произвольных скелетных мышц…
TRIVIA ВОПРОС: Как вы думаете, почему ваша походка меняется в состоянии алкогольного опьянения? Пьяные люди при ходьбе шатаются. Это почему? Влияет ли алкоголь на произвольные мышцы или нервную систему?
Закрытие
Теперь можно еще многое обсудить о мышечной системе, но это все, что мы пойдем в этом вводном видео.После просмотра этого видео вы должны хотя бы получить общее представление о том, из чего состоит мышечная система, ее свойства и функции.
Для более глубокого изучения мышечной системы ищите будущие видеоролики CTE, в которых мы будем освещать такие темы, как 14 основных групп мышц, а также такие темы, как распространенные заболевания и состояния, характерные для мышечной системы.
.