Рисунок мышечная клетка: Мышечная и нервная ткани — урок. Биология, Человек (8 класс).
Мышечная клетка (мышечное волокно). Строение
Мышечная клетка, хотя и обладает основными компонентами, присущими всем клеткам человеческого тела, ее необходимо рассмотреть детальнее.
Сразу следует, что мышечная клетка отличается от других клеток нашего тела. Основные различия приведены ниже:
- Мышечная клетка имеет многоядерное строение, причем ядра расположены на периферии клетки.
Ядра мышечных клеток не способны к делению, их функция сосредоточена в формировании информации для строения белковой молекулы.
Мышечная клетка, в своей оболочке имеет клетки-сателлиты, которые, в отличие от ядер, обладают способностью к делению и служат для восстановления наших мышц (например, после микротравм, полученных в ходе интенсивных тренировок). - Мышечная клетка наполнена сократительными структурами – миофибриллами. Это, своего рода, параллельно расположенные нити, общее количество которых в клетке может составлять порядка двух тысяч.
Назначение миофибрилл – стягивание мышечного волокна под действием нервного импульса.
Миофибрилла состоит из чередующихся поперечных полос темного и светлого цвета. Светлые участки способны уменьшать свою длину (до полного исчезновения) пропорционально силе сокращения миофибриллы, а при расслаблении мышцы – восстанавливают свою протяженность.
Миофибрилла включает огромное количество нитей двух белков: миозина и актина, которые располагаются вдоль миофибриллы. Причем, миозин – толстые нити, а актин – тонкие нити. Этим и объясняется светло-темное полосатое строение миофибриллы (темные полосы – миозин, светлые полосы – актин).
Каждая наша мышца состоит из пучков мышечных волокон (симпласта), которые представляют собой совокупность мышечных клеток продолговатой цилиндрической формы, края этих клеток сужены. В поперечном разрезе мышечная клетка выглядит так:
Как правило, мышечные клетки очень длинные (до 14 см) и тонкие (около 50 мкм). Обычно их длина равна длине отдельной мышцы.
Обычно их длина равна длине отдельной мышцы.
Мышечные клетки образуют пучки, из которых, собственно, и состоят наши мышцы.
Следует уяснить, что каждая мышечная клетка в таком пучке окружена соединительной тканью. В ней находятся лимфатические сосуды, кровеносные сосуды и нервные волокна.
Совокупность пучков мышечных клеток заключена в оболочке соединительной ткани. У основания мышцы, эта соединительная ткань образует сухожилия, посредством которых мышца крепится к кости.
Более наглядно данная структура показана на рисунке:
Таким образом, усилие, создаваемое нашими мышцами, через сухожилия передается костям скелета, в результате чего наши кости перемещаются относительно друг друга – осуществляется движения.
Но, что же заставляет наши мышцы сокращаться, как формируется это усилие и как передается в мышцу? На эти и другие вопросы Вы найдете ответы в статье Сокращение мышц. Принцип работы мышцы человека.
© Твой Тренинг
Материалы данной статьи охраняются законом о защите авторских прав. Копирование без указания ссылки на первоисточник и уведомления автора ЗАПРЕЩЕНО!
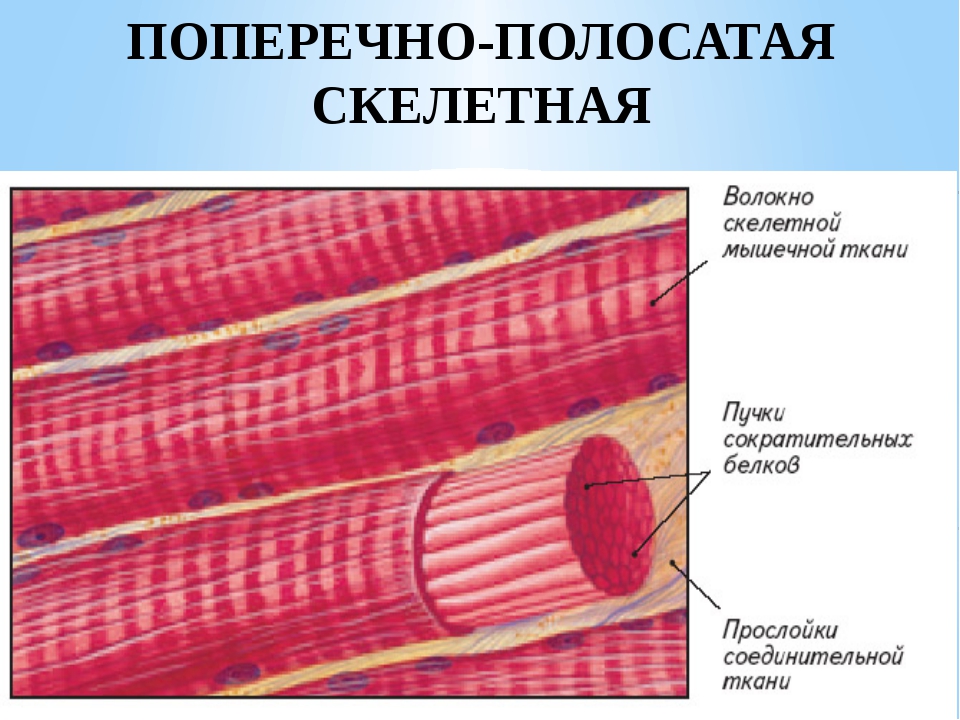
Поперечно-полосатая мышечная ткань
Мышечные
волокна – вытянутые в длину образования
цилиндрической формы, суживающиеся на
концах, покрытые оболочкой – сарколеммой.
Под сарколеммой в саркоплазме находятся
многочисленные ядра вытянутой по оси
волокна формы. Иногда ядра лежат попарно
или цепочкой, что указывает на их
способность к делению. Поперечно-полосатые
мышечные волокна имеют продольную и
поперечную исчерченность. Первая связана
с наличием в саркоплазме миофибрилл,
располагающихся вдоль мышечного волокна.
Поперечная исчерченность связана с
неоднородностью строения, различной
физико-химической организацией и разными
оптическими свойствами миофибрилл по
их длине.
Миофибрилла
состоит из чередующихся темных, способных
к двойному лучепреломлению участков –
анизотропных дисков и светлых, не
обладающих этой способностью
–
изотропных дисков. Миофибриллы относятся
к специальным органоидам и являются
морфологическим субстратом основной
специфической функции мышечной ткани
– сократимости.
Рис.
63. Поперечнополосатая мышечная ткань
Задание.
Рассмотреть микрофотографию (Рис. 63),
зарисовать его и сделать все необходимые
подписи к рисунку.
Гладкая мышечная ткань
Структурные
элементы гладкой мышечной ткани –
мышечные клетки. В продольном сечении
они веретенообразные, темноокрашенные
(Рис. 59). Их палочковидные ядра вытянуты
вдоль клеток. В поперечном сечении
мышечные клетки имеют форму округлых
ли многоугольных площадок различного
диаметра.
Рис.
64. Гладкая мышечная ткань: 1 – толстый
ядерный отдел клетки; 2 – заостренные
концы клетко; 3 – ядра; 4 – прослойки
базальной мембраны; 5 – прослойки
ареолярной ткани; 6 – сосуды; 7 – нервы;
8, 9 , 10, 11 – поперечные сечения мышечных
клеток; 12 – нервные клетки нервного
сплетения
Заостренные
концы одних мышечных клеток вклиниваются
между расширенными участками других,
формируя мышечный пласт, в котором
клетка контактирует со значительным
количеством соседних, объединяющихся
в «эффектор» –
моторную единицу.
Ядра
мышечных клеток имеют вытянутую форму
с глыбками хроматина и ядрышками. Форма
и структура ядер позволяет узнать
гладкую мышечную ткань, когда границы
отдельных клеток определить не удается.
Гладкие
мышечные клетки одеты обычной плазмалеммой
и базальной мембраной, ограничивающей
их от тончайших прослоек соединительной
ткани.
Рис.
65. Гладкая мышечная ткань
Задание.
Рассмотреть микрофотографию (Рис. 65),
65),
зарисовать его, сравнить с рисунком
(Рис. 64) и сделать все необходимые подписи
к нему.
Сердечная мышечная ткань
Миоциты
сердечной мускулатуры имеют структурные,
цитологические и функциональные
особенности (Рис. 66). Сердечные миоциты
в продольном сечении почти прямоугольные.
В центральной части клетки расположено
ядро овальной формы, вытянутое по оси.
В периферических отделах саркоплазмы
находятся пучки миофибрилл, обуславливающих
поперечную исчерченность.
Рис.
66. Сердечная мышечная ткань: 1- сердечные
миоциты; 2 – ядро, 3 — вставочные диски;
4 – соединительная ткань; 5 – капилляры
Характерным
морфологическим признаком сердечной
мышцы являются специфически организованные
контакты смежных миоцитов. Они выглядят
темными полосками и называются вставочными
дисками. Они образованы внутренними
листками сарколеммы соседних миоциов.
Таким
образом, с помощью вставочных дисков
сердечные миоциты объединяются в
мышечные комплексы, обеспечивающие
сокращение миокарда как единого целого.
Рис.
67. Сердечная мышечная ткань
Задание.
Рассмотреть микрофотографию (Рис. 67),
зарисовать его, сравнить с рисунком
(Рис. 66) и сделать все необходимые подписи
к нему.
Нервно-мышечная активность насекомых / Курс / Энтомология
Полёт, плавание, прыжки, ползание, сложнейшие акты строительной деятельности и брачного ритуала — всё разнообразие движений тела и его придатков, перистальтика кишечника, биение сердца и движения внутренних органов сводятся в конечном счёте к расслаблению и сокращению мышц, к преобразованию запасённой химической энергии в механическую.
Мышцы.
И скелетная, и висцеральная мускулатура насекомых образована поперечно-полосатыми волокнами, либо объединёнными в пучки мышц, либо распластанными по внутренним органам.
Обычно число волокон в каждой мышце невелико, но общее число мышц тела насекомых достигает 1 500, поэтому они намного превосходят по этому показателю более крупных млекопитающих. Впрочем, особенно мощные продольные крыловые мышцы у китайского дубового шелкопряда Antheraeapemyi G.-М. слагаются из 2 450 волокон толщиной около 45 мкм. Эти же мышцы у двукрылых образованы лишь шестью, но гораздо более толстыми волокнами (до 1 500 мкм в диаметре).
Впрочем, особенно мощные продольные крыловые мышцы у китайского дубового шелкопряда Antheraeapemyi G.-М. слагаются из 2 450 волокон толщиной около 45 мкм. Эти же мышцы у двукрылых образованы лишь шестью, но гораздо более толстыми волокнами (до 1 500 мкм в диаметре).
Прикрепление скелетных мышц насекомых к периодически сбрасываемым покровам обеспечивается проходящими сквозь клетки гиподермы тонофибриллами — тончайшими микротрубочками, связанными, с одной стороны, с десмосомами мышечных волокон, а с другой — с кутикулиновым слоем эпикугикулы (рис. 104).
Рисунок 104. Прикрепление скелетных мышц к покровам насекомых (по Romoser, 1981):
1 — эпикутикула; 2 — прокугикула; 3 — эпидермис; 4 — мышцы; 5 — кутикулин; 6 — поровые каналы; 7 — десмосомы; 8 — микротрубочка
В ходе линьки эти структуры замещаются новыми таким образом, что мышцы не теряют необходимые для их опоры связи. К тому же мембраны мышечных клеток прочно срастаются с базальной мембраной эпителия гиподермы.
Каждое мышечное волокно представляет собой видоизменённую глубоко специализированную клетку с собственной оболочкой — сарколеммой и многочисленными миофибриллами в саркоплазме (рис. 105). В этих клетках находится большое число ядер и особенно крупные митохондрии, что связывают с усиленным метаболизмом, а обилие элементов эндоплазматического (саркоплазматического) ретикулума — с необходимостью поглощения и выделения ионов Ca, активизирующих сократительные белки миофибрилл.
Рисунок 105. Схема организации мышечного волокна (по Romoser, 1981):
1 — саркомер; 2 — миофибрилла; 3 — саркоплазма; 4 — сарколемма
Весьма характерная для мышечных волокон насекомых система трансверзальных (поперечных) полых трубочек (Т-система), образованных глубокими впячиваниями сарколеммы, содействует переносу нервных импульсов на всю глубину саркоплазмы (рис. 106). Наиболее важные структуры мышечной клетки — миофибриллы — расположены плотными пакетами в саркоплазме, придают волокну характерную поперечную исчерченность и обусловливают его способность к сокращению. Исчерченность определяется чередованием светлых изотропных полос — 1-дисков — с более тёмными анизотропными А-дисками; сокращение обусловливают протофибриллы сократительных белков — актина и миозина (см. рис. 105).
106). Наиболее важные структуры мышечной клетки — миофибриллы — расположены плотными пакетами в саркоплазме, придают волокну характерную поперечную исчерченность и обусловливают его способность к сокращению. Исчерченность определяется чередованием светлых изотропных полос — 1-дисков — с более тёмными анизотропными А-дисками; сокращение обусловливают протофибриллы сократительных белков — актина и миозина (см. рис. 105).
Рисунок 106. Трубочки трансверзальной системы (Т-системы) мышц насекомых (по Smith, 1965)
Строение саркомера — участка миофибриллы, многократно повторяющегося по всей её длине, — типично для поперечно-полосатой мускулатуры как позвоночных животных, так и насекомых (рис.107). В соответствии с гипотезой скользящих нитей Хаксли более тонкие протофибриллы актина вдвигаются между толстыми протофибриллами миозина, что приводит к сокращению миофибриллы и мышечного волокна в целом. Полагают, что миозин, обладая свойствами АТФазы, трансформирует энергию макроэргических связей АТФ в механическую работу мускулатуры. Не вдаваясь в более подробное изложение этой гипотезы, ограничимся её демонстрацией (рис. 107).
Рисунок 107. Механизм мышечного сокращения — гипотеза скользящих нитей (по Тыщенко, 1977)
При сокращении обычных скелетных мышц длина их уменьшается на 25–30 %, но крыловые мышцы сокращаются лишь на 5–12 %. Однако последние сокращаются намного быстрее и с гораздо большей частотой, создавая необходимый для полёта ритм биения крыльев. В частности, многие бабочки и стрекозы совершают крыльями до 10–15 взмахов в секунду, жуки — до 175, пчелы — до 208–247, а мелкие двукрылые — до 500–1 000.
Столь необычный ритм сокращений недоступен мышцам позвоночных животных, сразу же сводимым судорогой и тетанусом при частоте возбуждений около 10 раз в секунду. Вместе с тем возникает проблема проведения нервных импульсов, в ответ на которые крыловые мышцы насекомых реагируют сокращениями. Распространяясь только по поверхности мембран, эти импульсы стимулируют выделение веществ, диффундирующих в саркоплазму, но даже самая высокая скорость диффузии все же недостаточна для последовательных стимуляций столь частых сокращений миофибрилл. Однако это противоречие устраняется развитием Т-системы, доносящей нервные импульсы до глубинных миофибрилл, а также уникальной способностью некоторых из них реагировать на один импульс серией частых сокращений.
Однако это противоречие устраняется развитием Т-системы, доносящей нервные импульсы до глубинных миофибрилл, а также уникальной способностью некоторых из них реагировать на один импульс серией частых сокращений.
Таким образом, мускулатура насекомых своеобразна, и это своеобразие, особенно явное для крыловых мышц, обусловлено несколькими типами волокон.
Трубчатые мышцы названы так потому, что в центре радиально расходящихся миофибрилл, как в полости трубки, заключены ядра (рис. 108, А). Способные к относительно медленным сокращениям, они обеспечивают движения ног насекомых, а также крыльев у стрекоз, тараканов и некоторых других форм. Реагируя дискретными сокращениями на каждый нервный импульс, трубчатые мышцы не требуют особенно больших доз кислорода и подходящие к ним трахеолы не проникают внутрь отдельных волокон.
Несколько более «быстрые» волокна плотноупакованных мышц отличаются положением ядер непосредственно под сарколеммой и заполнены плотными пачками миофибрилл (рис. 108, Б). Свойственные крыловой мускулатуре прямокрылых и чешуекрылых, они во всех прочих отношениях сравнимы с трубчатыми мышцами. Реагируя сокращениями на каждый нервный импульс, они вместе с последними именуются мышцами синхронного действия.
Быстрые фибриллярные мышцы образованы крупными миофибриллами с рассеянными между ними ядрами и митохондриями (рис.108, В). Характерные для звуковых органов цикад и крыловой мускулатуры жуков, перепончатокрылых и двукрылых, мышцы этого типа реагируют на один нервный импульс серией частых сокращений. По этому признаку они противопоставляются синхронным скелетным мышцам как мышцы асинхронного действия.
Рисунок 108. Строение трубчатых (А), плотноупакованных (Б) и фибриллярных (В) мышц насекомых (по Gillot, 1980):
1 — ядра; 2 — митохондрии
Скоростной машущий полет в сравнении с другими видами движения требует существенно больших затрат энергии. Летящая пчела расходует не менее 107 Дж/кг в час, что в 10 раз больше значения, рассчитанного для сердечной мышцы человека. Нуждаясь в усиленном обеспечении кислородом — в полете его расходование возрастает в 50 раз — фибриллярные мышцы снабжены проникающими внутрь волокон трахеолами и способны совершать работу, в десятки раз большую, нежели мышцы ног финиширующего спринтера. Во всяком случае, по скорости метаболизма фибриллярные крыловые мышцы намного превосходят все прочие ткани тела не только насекомых, но и животных других групп.
Нуждаясь в усиленном обеспечении кислородом — в полете его расходование возрастает в 50 раз — фибриллярные мышцы снабжены проникающими внутрь волокон трахеолами и способны совершать работу, в десятки раз большую, нежели мышцы ног финиширующего спринтера. Во всяком случае, по скорости метаболизма фибриллярные крыловые мышцы намного превосходят все прочие ткани тела не только насекомых, но и животных других групп.
В отличие от скелетной висцеральная мускулатура обеспечивает более медленные движения внутренних органов и не обладает глубокой специализацией. Сохраняя признаки поперечнополосатой мускулатуры, сократимые элементы висцеральных мышц обычно не сгруппированы в обособленные миофибриллы и часто вообще не иннервируются. Скелетные мышцы, напротив, иннервируются весьма обильно, формируя единые нервно-мышечные комплексы.
Проведение нервного возбуждения на мышцы.
Этот процесс связан со сложными электрохимическими явлениями на мембранах нервных клеток — нейронов. Изначально, то есть в исходном невозбуждённом состоянии, здесь регистрируется мембранный потенциал (МП), обусловленный неравновесной концентрацией ионов K+ по обе стороны мембраны и отрицательным зарядом цитоплазмы. В момент возбуждения сразу резко повышается проницаемость мембраны для ионов Na+, что приводит к падению МП и деполяризации данного участка нервного волокна. При этом он приобретает отрицательный заряд по отношению к соседним, невозбуждённым участкам, и регистрируемая между ними разность потенциалов (так называемый потенциал действия — ПД) индуцирует слабые токи, возбуждающие прилежащие участки. Таким образом, волна возбуждения переносится по нервному. волокну и скорость проведения нервного импульса достигает 0,3–25 м/с.
Вслед за падением МП проницаемость мембран для ионов Na+ снижается, но ионы K+ легко выводятся наружу. МП вновь повышается, однако для полного возвращения к исходному состоянию необходимо вывести наружу проникшие внутрь волокна ионы Na+ и заменить их ионами K+. Все эти функции выполняет «натриевый насос» мембраны, который, расходуя энергию, активно обменивает один ион Na+ на один ион K+.
Все эти функции выполняет «натриевый насос» мембраны, который, расходуя энергию, активно обменивает один ион Na+ на один ион K+.
Проведение возбуждения по нервам подчиняется некоторым правилам и ограничениям. В частности, ПД, начав распространяться в одном направлении, уже не меняет его впоследствии и сохраняет свою величину вне зависимости от силы раздражения или от пройденного пути. Дойдя до конца нервного волокна, нервный импульс переносится на мышцы и при этом вынужден преодолеть разделяющие их пространства.
В простейшем случае конец нерва образует тонкие варикозные разветвления на поверхности мышцы. Не сливающиеся с сарколеммой, они передают возбуждение мышцам через узкую зону контакта, не отграниченную от гемолимфы. В более сложном состоянии оболочка нерва полностью срастается с сарколеммой, изолируя область нервно-мышечного контакта (синапса) от гемолимфы (рис. 109). Под этой общей оболочкой сохраняется узкая (синаптическая) щель, препятствующая непосредственному переходу нервного импульса на мышечное волокно. Однако здесь же, в нервном окончании, содержатся многочисленные пузырьки (синаптические пузырьки), выделяющие специфические медиаторы возбуждения.
Рисунок 109. Схема нервно-мышечного синапса (по Тыщенко, 1977):
1 — мышечное волокно; 2 — ядро клетки; 3 — синаптическая щель; 4 — нейрилемма; 5 — митохондрии; 6 — синаптические пузырьки; 7 — миофибрилла
У позвоночных животных медиатором нервного возбуждения служит ацетилхолин, который, исполнив свои функции по переносу нервного импульса через синаптическую щель, сразу разрушается ферментом ацетил-холин-эстеразой. Оба эти соединения имеются и в нервной ткани насекомых, но есть веские основания считать медиатором возбуждения в нервно-мышечных контактах этих организмов глутаминовую кислоту, а медиатором торможения — амино-масляную кислоту.
Возбуждение мышечных волокон.
Нервный импульс, достигающий синаптической щели, содействует освобождению медиатора, который, в свою очередь, приводит к изменению проницаемости мембраны мышечного волокна. Отмеченное при этом падение мембранного потенциала содействует возникновению потенциала действия, распространяющегося по мышечному волокну и достигающего через Т-системы самых глубоких миофибрилл. Однако в отличие от проведения нервного импульса возбуждение мышечного волокна распространяется по всем возможным направлениям и в зависимости от силы раздражения постепенно затухает. При этом число сокращений синхронных мышечных волокон всегда совпадает с числом возникающих потенциалов, и если принять длительность одиночного сокращения с момента регистрации потенциала действия до расслабления мышцы за 30 мс, то за 1 с возможно осуществление не более 34 сокращений.
Таким образом, синхронная мускулатура не способна обеспечить высокий ритм сокращений крыловых мышц жуков, перепончатокрылых и двукрылых. Их асинхронные мышцы, реагируя на один импульс 4–13 биениями, позволяют развивать громадные для столь малых существ скорости полёта.
Иннервация мышечных волокон.
В отличие от мышц позвоночных животных мускулатура насекомых снабжена громадным числом нервных окончаний, причём чем «медленнее» мышца, тем этих окончаний больше. Например, в волокне межсегментной мышцы гусеницы Hyalophora cecropia L. отмечено до 1 000 синапсов, тогда как в каждом волокне разгибателя голени саранчи их не более 25. Столь богатая иннервация мышц насекомых, очевидно, содействует быстрому распространению возбуждения и обусловливает теснейшие связи с нервной системой в целом. Поэтому уместнее рассматривать комплексные нервно-мышечные единицы, которые у насекомых включают не менее 10–100 мышечных волокон и обслуживающий их нерв с 1–3 осевыми отростками (аксонами) нервных клеток (рис. 110). Один из них, именуемый «быстрым» аксоном, вызывает быстрые тетанические сокращения и расслабления мышц и наряду с «медленным» аксоном, ответственным за медленные тонические сокращения, служит пусковым нервным элементом. Кроме них в некоторых нервно-мышечных единицах присутствует тормозной регулирующий аксон. Таким образом, скелетная мускулатура насекомых находится под совместным контролем разнородных нервных центров.
Кроме них в некоторых нервно-мышечных единицах присутствует тормозной регулирующий аксон. Таким образом, скелетная мускулатура насекомых находится под совместным контролем разнородных нервных центров.
Рисунок 110. Нервно-мышечный комплекс (по Romoser, 1981):
1 — быстрый аксон; 2 — медленный аксон
Особенно своеобразна регуляция работы фибриллярных крыловых мышц. Способные к двукратному учащению ритма сокращений после ампутации крыльев, они характеризуются собственными миогенными водителями ритма в сочетании с более редкими нервными импульсами. Во всяком случае, даже изъятые из организма и убитые глицерином волокна фибриллярных мышц продолжают часами ритмично пульсировать в среде с АТФ и ионами Na+.
Tissue Triage and Freezing for Models of Skeletal Muscle Disease
Скелетных мышц является структурно и функционально уникальной ткани, и специализированные процедуры подготовки необходимы, чтобы позволить оптимальную оценку структурных и функциональных параметров. В то время как различные ткани, как правило, в замороженном патологических исследований в клинических и исследовательских контекстах, протоколы замораживания для не-мышечных тканей, как правило, связаны с полного погружения ткани в октябре перед замораживанием. Как показано на фиг.3, такой протокол не подходит для патологического оценки скелетных мышц и еще достаточно похож на протокол, описанный здесь, что это часто встречается ошибка. Целью данной работы является создание простой протокол для надлежащего обращения с мышцы, чтобы избежать проблем, как это. Консультации также был составлен на соответствующую обработки мышцы за пределы участка физиологических и клеточных исследований, с тем чтобы облегчить приобретение высококачественных данных по внешним основных лабораторий в случаях Wheповторно на месте исследования не являются предпочтительными или возможно.
Как отмечено в этом протоколе элементы, которые являются абсолютно необходимым в надлежащем обработки мышцы для патологических исследований включают минимизации содержание воды в тканях, снижение температуры, при которой мышца заморожены, и увеличить скорость, при которой замораживание достигается. Повышенная влажность в ткани или чрезмерной медлительности процесса замораживания (производства недостаточных температурах или отсутствием прямого контакта между агентом замерзания и ткани, как встречается с жидким азотом) приведет к замораживанию артефактов, которые могут повлиять на патологический анализ. Как октября обеспечивает дополнительный источник увлажнением тканей, многие лаборатории использовать другие клеи, как трагакантовая как вложения подложки. Даже в тех случаях, когда замораживание выполняется надлежащим образом, следует позаботиться, чтобы избежать впоследствии случайно оттаивания образцов при контакте с RT контейнерах или инструментов.Таким образом, успешный процесс замораживания требует определенной планирования, которая включает предварительно охлаждение всех инструментов и контейнеров, которые будут использоваться. Когда артефакты морозильные встречаются, существует метод, описанный здесь для восстановления тканей, которое является достаточным для большинства патологических оценок. Однако это замораживания / оттаивания не предлагает идеальный гистологию и имеет потенциал, чтобы нарушить другие молекулярные или ферментативные исследования ткани (в дополнение к времени, необходимого для повторного замораживания ткани), так с использованием соответствующих исходных практики замораживания является гораздо предпочтительнее . В случаях, когда замораживание артефакт, возникающих на предварительно замороженной ткани, однако, способ, описанный здесь, может быть чрезвычайно полезным.
Повышенная влажность в ткани или чрезмерной медлительности процесса замораживания (производства недостаточных температурах или отсутствием прямого контакта между агентом замерзания и ткани, как встречается с жидким азотом) приведет к замораживанию артефактов, которые могут повлиять на патологический анализ. Как октября обеспечивает дополнительный источник увлажнением тканей, многие лаборатории использовать другие клеи, как трагакантовая как вложения подложки. Даже в тех случаях, когда замораживание выполняется надлежащим образом, следует позаботиться, чтобы избежать впоследствии случайно оттаивания образцов при контакте с RT контейнерах или инструментов.Таким образом, успешный процесс замораживания требует определенной планирования, которая включает предварительно охлаждение всех инструментов и контейнеров, которые будут использоваться. Когда артефакты морозильные встречаются, существует метод, описанный здесь для восстановления тканей, которое является достаточным для большинства патологических оценок. Однако это замораживания / оттаивания не предлагает идеальный гистологию и имеет потенциал, чтобы нарушить другие молекулярные или ферментативные исследования ткани (в дополнение к времени, необходимого для повторного замораживания ткани), так с использованием соответствующих исходных практики замораживания является гораздо предпочтительнее . В случаях, когда замораживание артефакт, возникающих на предварительно замороженной ткани, однако, способ, описанный здесь, может быть чрезвычайно полезным.
Фиксация и обработка ткани для ЭМ может предложить конкретные технические проблемы, которые требуют планирования до сбора ткани. При сборе образцов для EM наиболее распространенная ошибка предполагает использование тканевых фрагментов, которые являются слишком толстым для glutaraldehydе проникнуть. Как глютаральдегид только проникает около 0,1 см в мышечную ткань от заданной поверхности, следует позаботиться, чтобы убедиться, что одно измерение образцов ЭМ является не толще 0,2 см.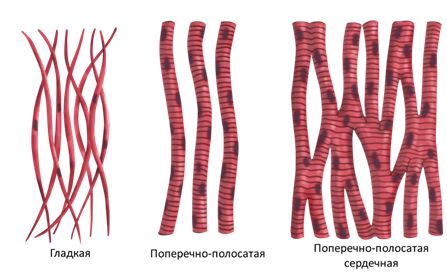 Кроме того, как EM является отличным средством непосредственно оценки сократительной аппарата, некоторые исследователи разработали стратегии для предварительного натяжения или предварительного натяжения мышц до фиксации, чтобы позволить измерение сократительных элементов в физиологически соответствующего напряжения. Там нет стандартного протокола для предварительного натяжения, но две стратегии кратко описаны в этом протоколе. Следует отметить, что попытки предварительного натяжения мышц может привести к непредсказуемым результатам, если они не сделаны в очень специфическим образом, и это может быть предпочтительным, чтобы исправить мышцы в ослабленном состоянии, чтобы предотвратить артефактом изменения длины саркомера через нестандартные предварительного натяжения процедура 8,9. Для мышц, в которых такие специфические измерения не нужны (в том числе большей Ъiopsies выполненные для клинических целей), усилия по предварительному натяжению мышц, как правило, не производится, и основной эффект на мышечную морфологии является неравномерное расстояние саркомеров в мышцах.
Кроме того, как EM является отличным средством непосредственно оценки сократительной аппарата, некоторые исследователи разработали стратегии для предварительного натяжения или предварительного натяжения мышц до фиксации, чтобы позволить измерение сократительных элементов в физиологически соответствующего напряжения. Там нет стандартного протокола для предварительного натяжения, но две стратегии кратко описаны в этом протоколе. Следует отметить, что попытки предварительного натяжения мышц может привести к непредсказуемым результатам, если они не сделаны в очень специфическим образом, и это может быть предпочтительным, чтобы исправить мышцы в ослабленном состоянии, чтобы предотвратить артефактом изменения длины саркомера через нестандартные предварительного натяжения процедура 8,9. Для мышц, в которых такие специфические измерения не нужны (в том числе большей Ъiopsies выполненные для клинических целей), усилия по предварительному натяжению мышц, как правило, не производится, и основной эффект на мышечную морфологии является неравномерное расстояние саркомеров в мышцах.
Эта статья представляет собой первый в серии, чтобы обеспечить СОП для проведения испытаний в области врожденного заболевания мышц, и это являет собой результат труда более чем 20 специалистов в врожденной мышечной области заболевания, которые обычно выполняют клеточном, молекулярном, функциональный, физиологические и патологическое исследование. Диапазон опубликованных СОП будут доступны в течение следующего года, и были обсуждены необходимые протоколы и соответствующие форматы публикации для каждого при врожденном мышц Болезнь Консорциум семинара, проведенного в апреле 2013 года в Вашингтоне, округ Колумбия Цель этой SOP усилий является обеспечение дорожная карта необходимого тестирования и анализа образцов в врожденной области заболевания мышц, чтобы 1) стандартизации практики и конечные точки, используемые в нашей области, как муч насколько возможно, и 2) обеспечить обучение на стандартной практикой для новых исследователей в нашей области. Мы считаем, что эти ресурсы будут способствовать появлению новых исследователей в нашей под изученный поле и тем самым улучшить сферу исследований, которые могут быть выполнены. Кроме того, стандартизация практики будет чрезвычайно полезно сравнить данные по исследованиям и определить конечные точки при планировании и выполнении доклинические и клинические испытания.
Мы считаем, что эти ресурсы будут способствовать появлению новых исследователей в нашей под изученный поле и тем самым улучшить сферу исследований, которые могут быть выполнены. Кроме того, стандартизация практики будет чрезвычайно полезно сравнить данные по исследованиям и определить конечные точки при планировании и выполнении доклинические и клинические испытания.
В то время как основное внимание в этой статье связано с соответствующей замораживания и приготовления ткани для различных исследований, наша совместная группа также обсудила полезные конечные точки для патологического анализа мышечных образцов. В настоящее время нет официального консенсуса на подходе принять при выполнении патологический анализ, и множество различных исследований выполнено связать новые данные с предыдущими публикаций для каждого соответствующего заболевания. Таким образом, мы думали, что было бы полезно, чтобыпредложить некоторые общие рекомендации по планированию патологических конечных точек в мышечной патологии характеристики. До количественного патологию в исследовании, значительная мысль должна быть введена в 1) метод измерения размера волокно, 2) возможность аномалий волоконно-типоспецифических или эффектов лечения, 3) возможность аномалий или эффектов, которые ограничены в отдельных мышц, и 4) стратегия для количественного патологических, которые характерны для этой болезни в исследовании. Размер волокна является необходимым конечная точка для большинства исследований, и, к сожалению, весьма обширен изменение в том, как она количественно. Многие исследования показывают эти результаты, используя автоматизированные методы количественной предоставляемые проприетарного программного обеспечения визуализации, но многие из этих программ срезать углы (например, если предположить, что волокна представляют собой окружности или эллипсы), которые могут сделать эти автоматизированные измерения неточны. Надо понять, как эти автоматизированные программы делают их измерения бEfore имея уверенность в измерениях, и мы призываем следователей включить эти данные в методах бумаги. Кроме того, удельный измерение используется для обозначения размера волокна является чрезвычайно переменным, и некоторые измерения предпочтительнее других 10,11. Обычно используется измерение размера волокон является площадь поперечного сечения волокна (CSA), в частности, потому, что исследователи, выполняющие физиологических исследований стандартизировать свои результаты измерений, полученных с помощью CSA свои инструменты. К сожалению, в то время как измерения CSA может точно отражать размер волокна в идеальных секций поперечными, они широко в зависимости от ориентации волокон (в том случае, продольные или наклонно-секционные слои будут иметь искусственно завышенные размеры CSA) и, таким образом, не идеальными для измерения размера волокон. Предпочтительным измерение размера волокон, которые меньше зависит от волокна площади поперечного сечения является минимальной диаметр Фере (диаметр MinFeret), который является измерение гое малый диаметр в мышечной клетке 12. Это измерение слабо зависит от ориентации волокон и, как правило, клиническая золотым стандартом для измерения волокна, и следователи поощряются двигаться к использованию этой техники. Эти измерения часто можно сделать с помощью того же программного обеспечения, который генерирует измерений CSA 13, и также просто для измерения вручную. Что касается оценки патологических данных в зависимости от типа волокна, определенной мышцы, и в контексте патологических, связанных с конкретным заболеванием, они менее спорные вопросы, которые должны просто быть рассмотрены при планировании исследования. Тип волокна можно оценить с помощью иммуногистохимического или АТФазную окрашивание, но это полезно рассмотреть, что конкретные мышцы и животных имеют особые смеси этих типов волокон (что требует разные ожидания и тестирование стратегии). Мышцы конкретных патологических участие или эффективность леченияможет иметь место, и общий вес мышц по сравнению с контрольной может быть использован, чтобы идентифицировать степень гетерогенности заболевания, прежде чем решить на мышцы к патологически оценки.
Кроме того, удельный измерение используется для обозначения размера волокна является чрезвычайно переменным, и некоторые измерения предпочтительнее других 10,11. Обычно используется измерение размера волокон является площадь поперечного сечения волокна (CSA), в частности, потому, что исследователи, выполняющие физиологических исследований стандартизировать свои результаты измерений, полученных с помощью CSA свои инструменты. К сожалению, в то время как измерения CSA может точно отражать размер волокна в идеальных секций поперечными, они широко в зависимости от ориентации волокон (в том случае, продольные или наклонно-секционные слои будут иметь искусственно завышенные размеры CSA) и, таким образом, не идеальными для измерения размера волокон. Предпочтительным измерение размера волокон, которые меньше зависит от волокна площади поперечного сечения является минимальной диаметр Фере (диаметр MinFeret), который является измерение гое малый диаметр в мышечной клетке 12. Это измерение слабо зависит от ориентации волокон и, как правило, клиническая золотым стандартом для измерения волокна, и следователи поощряются двигаться к использованию этой техники. Эти измерения часто можно сделать с помощью того же программного обеспечения, который генерирует измерений CSA 13, и также просто для измерения вручную. Что касается оценки патологических данных в зависимости от типа волокна, определенной мышцы, и в контексте патологических, связанных с конкретным заболеванием, они менее спорные вопросы, которые должны просто быть рассмотрены при планировании исследования. Тип волокна можно оценить с помощью иммуногистохимического или АТФазную окрашивание, но это полезно рассмотреть, что конкретные мышцы и животных имеют особые смеси этих типов волокон (что требует разные ожидания и тестирование стратегии). Мышцы конкретных патологических участие или эффективность леченияможет иметь место, и общий вес мышц по сравнению с контрольной может быть использован, чтобы идентифицировать степень гетерогенности заболевания, прежде чем решить на мышцы к патологически оценки. Наконец, хорошо известно, что многие заболевания мышц, связанные с конкретными патологических отклонений (например, немалиновой стержней в немалиновой миопатия) 14,15, и поэтому он также полезно рассмотреть, являются ли эти нарушения обнаружены в волоконно-типа или мышцы конкретных распределение при выполнении анализа 16,17. В целом, в то время как мы не предлагаем негибкий набор стандартов для оценки мышцы, мы считаем, что эти вопросы должны рассматриваться до исполнения патологических исследований в любой скелетной мышечных заболеваний.
Наконец, хорошо известно, что многие заболевания мышц, связанные с конкретными патологических отклонений (например, немалиновой стержней в немалиновой миопатия) 14,15, и поэтому он также полезно рассмотреть, являются ли эти нарушения обнаружены в волоконно-типа или мышцы конкретных распределение при выполнении анализа 16,17. В целом, в то время как мы не предлагаем негибкий набор стандартов для оценки мышцы, мы считаем, что эти вопросы должны рассматриваться до исполнения патологических исследований в любой скелетной мышечных заболеваний.
Участие мезенхимных стромальных клеток в регенерации мышечной ткани • О. В. Паюшина, Е. И. Домарацкая, О. Н. Шевелева • Журнал общей биологии • Выпуск 1 • Том 80, 2019 г.
Клеточная терапия с использованием мезенхимных стромальных клеток (МСК) является перспективным подходом к восстановлению скелетных мышц после травм и заболеваний. МСК, исходно присутствующие в мышце или приходящие в нее из костного мозга в ответ на повреждение, вырабатывают различные регуляторные молекулы, которые благотворно влияют на выживание, размножение и дифференцировку клеток, усиливают рост кровеносных сосудов, оказывают противовоспалительное и антифибротическое действие. Способность к продукции разнообразных факторов, воздействующих на все стадии репаративного процесса, позволяет использовать МСК для ускорения регенерации, что было неоднократно показано на различных экспериментальных моделях повреждения мышц. Эффект МСК может быть усилен путем совершенствования способов доставки клеток в ткань и улучшения их выживаемости; кроме того, секреторный профиль клеток может быть изменен в необходимом направлении путем воздействия различных физических или химических стимулов либо с помощью генетической модификации. Новым направлением регенеративной медицины становится использование продуцируемых МСК внеклеточных везикул и содержащихся в них регуляторных молекул, прежде всего микроРНК. Активацию регенеративного потенциала МСК можно рассматривать как инструмент тканевой инженерии in vivo, стимулирующий восстановление тканей за счет внутренних резервов.
Повреждение скелетных мышц такими воздействиями, как сдавливание, порезы, проколы, отморожения, истощающие физические нагрузки, может приводить к образованию рубцов и значительно ухудшать сократительную способность. Существуют разные подходы к восстановлению мышц после травм и заболеваний, и один из наиболее перспективных связан с использованием мезенхимных стромальных клеток (МСК) (Заглавная иллюстрация). МСК привлекательны для клеточной терапии своей доступностью, легкостью культивирования, несклонностью к образованию опухолей и способностью не отторгаться иммунной системой. Они способны давать начало различным типам зрелых клеток и выделять множество регуляторных молекул, оказывающих благотворное влияние на ткань. В скелетных мышцах, как и во многих других тканях и органах, присутствуют клетки со свойствами МСК, которые в случае травмы размножаются и участвуют в регенерации. Строго говоря, главная роль в восстановлении мышц принадлежит не им, а миосателлитам – незрелым предшественникам, способным дифференцироваться в клетки, сливающиеся затем в мышечные волокна. Однако вклад МСК и близких к ним клеточных популяций, содержащихся в мышцах, также не стоит недооценивать: они создают временную нишу для мышечных предшественников, секретируя молекулы, способствующие их делению и дифференцировке. Кроме того, при травме мышцы в нее приходят МСК из костного мозга. Их привлекают вещества, так называемые хемоаттрактанты, выделяемые тканью в условиях нехватки кислорода из-за повреждения. Эти пришлые клетки тоже участвуют в восстановительном процессе наряду с местными МСК мышц.
Каковы же механизмы участия МСК в регенерации мышечной ткани? Эксперименты на клеточных культурах и на животных с поврежденными мышцами показали, что МСК способны непосредственно давать начало мышечным клеткам или сливаться с ними. Однако такие события происходят редко, и их вклад в регенерации едва ли может быть существенным. Гораздо более важную роль в ней играет способность МСК секретировать биологически активные вещества, способствующие улучшению состояния поврежденной ткани. Эти вещества предотвращают гибель мышечных клеток, стимулируют их размножение и слияние, уменьшают воспаление, усиливают рост сосудов, а значит, и кровоснабжение регенерирующей мышцы. По некоторым данным, они также препятствуют образованию рубца на месте травмы. Таким образом, выделяемые МСК факторы воздействуют на все стадии восстановительного процесса. А значит, дополнительное введение МСК в поврежденную мышцу можно рассматривать как способ комплексной доставки в нее биоактивных молекул для ускорения регенерации.
Эти вещества предотвращают гибель мышечных клеток, стимулируют их размножение и слияние, уменьшают воспаление, усиливают рост сосудов, а значит, и кровоснабжение регенерирующей мышцы. По некоторым данным, они также препятствуют образованию рубца на месте травмы. Таким образом, выделяемые МСК факторы воздействуют на все стадии восстановительного процесса. А значит, дополнительное введение МСК в поврежденную мышцу можно рассматривать как способ комплексной доставки в нее биоактивных молекул для ускорения регенерации.
Этот подход показал свою эффективность в многочисленных опытах на животных. Мышцы мышей или крыс травмировали сдавливанием или надрезанием, разрушали инъекциями токсинов, вызывали мышечную атрофию с помощью высокожировой диеты или длительной иммобилизации конечности, нарушали кровоток в мышцах, перевязывая бедренную артерию, использовали мутантных животных с генетически обусловленной миодистрофией. И во всех этих случаях введение МСК приводило к улучшению структуры поврежденных мышц и их функционального состояния.
Таким образом, результаты использования МСК для регенерации мышц выглядят весьма обнадеживающими. Но эффективность их применения можно сделать еще более высокой, если усовершенствовать доставку клеток в поврежденную ткань, улучшить их выживаемость в организме реципиента или усилить продукцию ими биоактивных молекул. В частности, высокой концентрации МСК в месте повреждения можно достичь, трансплантируя их на искусственных носителях. Разрабатывается и другой подход: на мышцу воздействуют импульсами фокусированного ультразвука, что способствует усиленной миграции в нее МСК. Известна также магнитная система доставки МСК, когда вводимые клетки метят наночастицами оксида железа, а к поврежденной области прикладывают внешнее магнитное поле. В этом случае содержание донорских клеток в травмированной мышце оказывается выше, а ее восстановление эффективнее, чем после трансплантации МСК обычным образом. Еще один путь к повышению регенеративного потенциала МСК состоит в воздействии на них различными физическими или химическими факторами (сниженным содержанием кислорода, механическим натяжением, гормонами, фармакологическими агентами и т.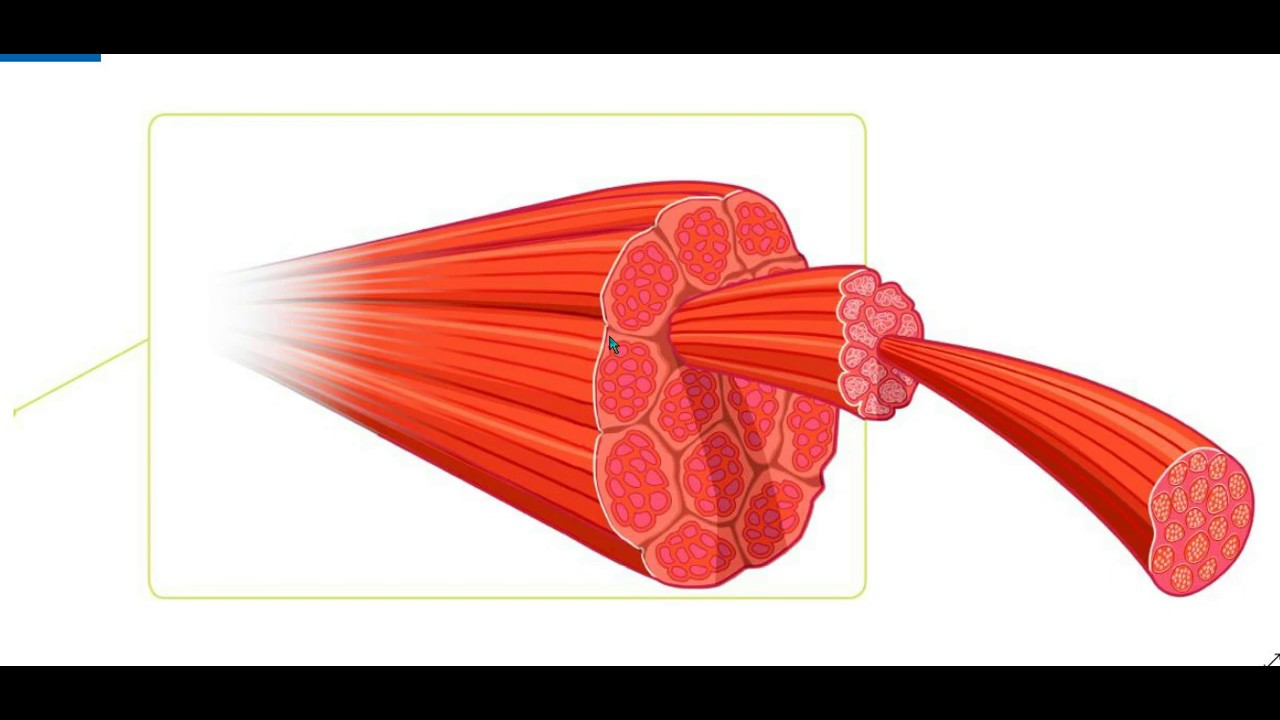 п.), стимулирующими их секреторную активность. Таким образом можно добиться усиленного выделения клетками тех молекул, которые необходимы для регенерации. С той же целью можно подвергнуть МСК генетической модификации, введя в них гены, обеспечивающие синтез необходимых веществ. Наконец, вводить в поврежденную ткань можно не сами МСК, а их внеклеточные везикулы – мембранные пузырьки, заключающие в себе различные регуляторные молекулы. Эти везикулы могут стимулировать клетку-мишень через мембранные рецепторы или, сливаясь с ней, доставлять свое содержимое в цитоплазму.
п.), стимулирующими их секреторную активность. Таким образом можно добиться усиленного выделения клетками тех молекул, которые необходимы для регенерации. С той же целью можно подвергнуть МСК генетической модификации, введя в них гены, обеспечивающие синтез необходимых веществ. Наконец, вводить в поврежденную ткань можно не сами МСК, а их внеклеточные везикулы – мембранные пузырьки, заключающие в себе различные регуляторные молекулы. Эти везикулы могут стимулировать клетку-мишень через мембранные рецепторы или, сливаясь с ней, доставлять свое содержимое в цитоплазму.
В последние годы получение МСК с заданными свойствами, необходимыми для полноценного восстановления тканей, становится одной из активно разрабатываемых областей регенеративной медицины. В перспективе знание биологии этих клеток позволит управлять их миграцией к месту повреждения и влиять на их секреторный профиль, что обеспечит эффективную тканевую регенерацию за счет внутренних резервов.
Мышечная система человека. Подготовка к огэ биология.
Мышечная система человека
ОГЭ биология
1. В состав какого органа входят мышечные клетки, изображённые на рисунке?
1) кишечник
2) двуглавая мышца плеча
3) матка
4) Желудок
2. В состав какого органа входят мышечные клетки,
изображённые на рисунке?
2) кишечник
3) матка
4) Желудок
3. В состав какого органа входят мышечные клетки, изображённые на рисунке?
1) языка
2) двуглавой мышцы
3) височной мышцы
4) стенки желудка
4. На рисунке изображены бицепс (1) и трицепс (2). Что
произойдёт с этими мышцами, если согнуть руку в локте?
1) Бицепс сократится, а трицепс расслабится.
2) Бицепс сократится, а трицепс не изменится.
3) Трицепс сократится, а бицепс расслабится.
4) Трицепс сократится, а бицепс не изменится.
5. Какой цифрой на рисунке обозначен бицепс?
6. Какой признак свидетельствует о физической
Какой признак свидетельствует о физической
тренированности человека?
1) нормальное артериальное давление
2) диаметр бицепса
3) умение концентрироваться при выполнении работы
4) нормальная масса тела
7. Верны ли следующие суждения о мышечных тканях человека?
А. Поперечнополосатая скелетная мышечная ткань состоит из одноядерных веретеновидных клеток.
Б. Гладкая мышечная ткань сокращается медленно
и непроизвольно.
1) верно только А
2) верно только Б
3) верны оба суждения
4) оба суждения неверны
8. Какой цифрой на рисунке обозначены мышцы, сгибающие
и вращающие туловище вокруг продольной оси?
9. На рисунке изображены бицепс (1) и трицепс (2). Что
произойдёт с этими мышцами, если разогнуть руку в локте?
1) Бицепс сократится, а трицепс расслабится.
2) Бицепс сократится, а трицепс не изменится.
3) Трицепс сократится, а бицепс расслабится.
4) Трицепс сократится, а бицепс не изменится.
10. Какой цифрой на рисунке обозначены мимические
мышцы?
11. Какой цифрой на рисунке обозначен трицепс?
12. Производительность физического труда будет выше, а утомление наступит позже, если работу выполнять в
1) медленном ритме и с малой нагрузкой
2) среднем ритме и со средней нагрузкой
3) среднем ритме с большой физической нагрузкой
4) в медленном ритме с большой физической нагрузкой
13. Каким номером на рисунке обозначено
мышечное волокно?
14. Для возникновения мышечного
сокращения необходимы ионы
- Калия 2) кальция
3) железа 4) магния
15. В развитии утомления работающих мышц у человека ведущую роль играет
1) истощение запаса АТФ в организме в процессе выполнения работы
2) время начала работы и её продолжительность
3) нарастание усталости в самих мышцах при работе
4) торможение нервных центров, регулирующих работу мышц
16. Координация работы мышц-разгибателей осуществляется в
Координация работы мышц-разгибателей осуществляется в
1) сухожилиях
2) рецепторах
3) нервных центрах
4) скелетных мышцах
17. Какой цифрой на рисунке обозначены
мышцы туловища?
18. Какая мышца легко прощупывается в области плеча при согнутой в локте руке?
1) зубчатая мышца
2) сгибатель пальцев
3) двуглавая мышца
4) портняжная мышца
19. Рассмотрите рисунок. В каком состоянии находятся нервные центры двуглавой и
трёхглавой мышц в момент сгибания руки?
- Нервный центр двуглавой мышцы возбуждён,
а трёхглавой заторможен.
2) Нервный центр двуглавой мышцы заторможен,
а трёхглавой возбуждён.
3) Нервные центры мышц заторможены.
4) Нервные центры мышц возбуждены.
20. Рассмотрите рисунок. В каком состоянии находятся нервные центры двуглавой и трёхглавой мышц в процессе разгибания руки?
1) Нервный центр двуглавой мышцы возбуждён, а трёхглавой заторможен.
2) Нервный центр двуглавой мышцы заторможен, а трёхглавой возбуждён.
3) Нервные центры мышц заторможены.
4) Нервные центры мышц возбуждены.
21. В каком состоянии должны находиться нервные центры
головного мозга, отвечающие за сгибание и разгибание
руки, чтобы человек мог удержать предмет на
вытянутой руке?
1) оба центра расслаблены
2) оба центра возбуждены
3) центр сгибания возбуждён, а разгибания расслаблен
4) центр разгибания возбуждён, а сгибания расслаблен
22. Штангисты и культуристы стремятся к тому, чтобы
1) увеличить площадь поперечного сечения мышц
2) увеличить длину мышц
3) увеличить частоту сокращений мышц
4) снизить частоту сокращений мышц
23. В приведённой ниже таблице между позициями первого и второго столбца имеется взаимосвязь.
Объект Процесс
Нейрон Проведение
… Сокращение
Какое понятие следует вписать на место пропуска в этой таблице?
1) лимфоцит
2) эпителиоцит
3) эритроцит
4) Миоцит
24. В приведённой ниже таблице между позициями первого и второго столбца имеется взаимосвязь.
В приведённой ниже таблице между позициями первого и второго столбца имеется взаимосвязь.
Объект Процесс
Нейрон Проведение
… Сокращение
Какое понятие следует вписать на место пропуска в этой таблице?
1) лизосома
2) мышечное волокно
3) эпителий
4) соединительная ткань
25. К поперечнополосатой мускулатуре относятся
- мышцы стенок кишечника
2) мышцы стенок артерий
3) мышцы стенок желудка
4) мышцы, сгибающие руку
26. Между биологическими объектами и процессами, указанными в столбцах приведённой ниже таблицы, имеется определённая связь.
ОБЪЕКТ ПРОЦЕСС
соединительная ткань кость
… миокард
Какое понятие следует вписать на место пропуска в этой таблице?
1) мышечная ткань 2) нервная ткань
3) покровная ткань 4) железистая ткань
27. К гладкой мускулатуре относятся
- мышцы стенок сердца
2) мышцы стенок артерий
3) мышцы, разгибающие ногу
4) мышцы, сгибающие руку
Задание 4 ЕГЭ по биологии 2021: теория и практика
РНК (рибонуклеиновая кислота) – одна полинуклеотидная цепь нуклеотидов (может иметь прямые и спиральные участки, образовывать петли). Масса РНК в несколько сот раз меньше чем масса ДНК
РИБОнуклеотид:
1. Один из четырех типов азотистых оснований:
аденин (А), гуанин (Г), цитозин (Ц) и урацил (У)
2. Пятиуглеродный моносахарид (углевод) — рибоза
3. Молекула фосфорной кислоты
Функции РНК:
1. РНК — хранитель генетической информации у ретровирусов, выполняют у них функцию хромосом
2. Информационная/матричная (и-РНК/м-РНК) – 5% от всех РНК клетки — «копирует» и переносит информацию о первичной структуре белка (последовательности аминокислот) и следовательно функциях белков от ДНК к месту синтеза белков на рибосомах в цитоплазме
3. Транспортная (т-РНК) – 10% от всех РНК клетки — переносит аминокислоты из цитоплазмы к месту синтеза белка на рибосомы и осуществляет точную ориентацию аминокислоты (по принципу комплементарности)
4. Рибосомная (р-РНК) – 85% от всех РНК клетки — синтезируется на участках ДНК хромосом, расположенных в ядрышке, а содержится в рибосомах. В комплексе с белками образует рибосомы (органоиды клетки, на которых синтезируются белки)
Рибосомная (р-РНК) – 85% от всех РНК клетки — синтезируется на участках ДНК хромосом, расположенных в ядрышке, а содержится в рибосомах. В комплексе с белками образует рибосомы (органоиды клетки, на которых синтезируются белки)
ДНК (дезоксирибонуклеиновая кислота) – две спиральные полинуклеотидные цепи, соединенные друг с другом водородными связями и состоящие из десятков тысяч или миллионов мономеров
ДЕЗОКСИнуклеотид:
1. Один из четырех типов азотистых оснований: аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т)
2. Пятиуглеродный моносахарид (углевод) – дезоксирибоза
3. Молекула фосфорной кислоты
Функции ДНК:
1. Хранение наследственной информации (в форме хроматина – деспирализованная цепь ДНК в комплексе с белками-гистонами, после спирализации образует хромосомы; наследственная информация закодирована в генетическом коде – последовательности нуклеотидов)
2. Воспроизведение наследственной информации (биосинтез белка (транскрипция и трансляция) – это и есть воспроизведение генетической информации). Последовательность нуклеотидов ДНК определяет (кодирует) первичную структуру белков организма
3. Передача наследственной информации следующим поколениям (при делении клеток происходит репликация ДНК, так, что у дочерних клеток получаются копии генетического материала). Эти последовательности индивидуальны и для каждого вида организмов и для отдельных особей
Как нарисовать мышечные клетки
Мышечная ткань состоит из мышечных клеток. Мышечные клетки проявляют три основных свойства, таких как возбудимость, проводимость и сократимость. Мышечные клетки имеют удлиненную форму волокон (миоцитов). Миоциты имеют несколько миофибрилл, которые отвечают за чередование темных и светлых полос на поперечно-полосатых или полосатых мышцах. Скелетная мышца состоит из полосатых мышечных клеток. Каждое волокно скелетных мышц представляет собой длинную цилиндрическую неразветвленную клетку.Это многоядерная клетка с множеством овальных ядер. Эти клетки работают под сознательным контролем организма.
Эти клетки работают под сознательным контролем организма.
Полосатая мышечная клетка имеет форму длинного волокна, поэтому мы не изображаем концы этой клетки.
Нарисуйте две параллельные кривые и отметьте несколько темных полос на одинаковом расстоянии, как показано.
Сделайте овальные ядра и этикетку.
Гладкая мускулатура расположена в стенках внутренних органов, таких как кровеносные сосуды, трахея, бронхи, желудок, кишечник, выводные и половые протоки и т. Д.Эти мышечные клетки также присутствуют в радужной оболочке глаза и цилиарном теле глаза и в коже в виде арректорных пилей, прикрепленных к волосяным фолликулам. Обычно эти ячейки располагаются в виде листов. Гладкомышечное волокно веретенообразное, одноядерное. Миофибриллы не показывают чередующихся темных и светлых полос из-за неправильного расположения молекул актина и миозина. Они не работают под сознательным контролем. Они могут длительное время оставаться в сжатом состоянии без усталости. Они находятся под контролем автономной нервной системы.
Нарисуйте форму веретена и несколько кривых, представляющих миофибриллы. В центре нарисуйте одно круглое ядро.
Сердечная мышца имеет поперечно-полосатую форму, как и скелетная мышца. Он находится в миокарде сердца позвоночных. Эти клетки представляют собой короткие цилиндрические, одноядерные или двуядерные клетки, концы которых разветвляются и образуют соединения с другими клетками сердечной мышцы. Темные линии на клетках сердечной мышцы называются интеркалированными дисками.
Нарисуйте четыре или пять рядов полосатых мышечных клеток и случайным образом соедините их ветвями.
Нарисуйте интеркалированные диски и отметьте несколько полос.
Наконец, нарисуйте ядра, как показано.
Схематический рисунок небольшого среза мышечной клетки, показывающий некоторые …
Контекст 1
… Сокращение мышц является результатом силовых взаимодействий между поперечными мостиками миозина на толстой нити и актином на тонкой нити . Силовые взаимодействия регулируются Ca 2+ через специализированные белки тонкой нити. Это является спорным, как сократительный и регуляторных систем динамически взаимодействуют, чтобы определить ход времени мышечного сокращения и расслабления.В то время как кинетику Ca 2+ -индуцированной регуляции тонких филаментов часто исследуют с использованием изолированных белков, кинетику силы обычно изучают в мышечных волокнах. Разрыв между исследованиями изолированных белков и структурированных волокон в настоящее время устраняется с помощью новейших методов, которые анализируют химическую и механическую кинетику небольших компонентов мышечного волокна, субклеточных миофибрилл, изолированных из скелетных и сердечных мышц. Сформированные из последовательно расположенных повторяющихся единиц, называемых саркомерами, миофибриллы имеют полный полностью структурированный ансамбль сократительных и регуляторных белков Ca 2+.Малый диаметр миофибрилл (несколько микрометров) облегчает анализ кинетики сокращения и расслабления саркомера, вызванных быстрыми изменениями [АТФ] или [Ca 2+]. Среди процессов, изучаемых на миофибриллах: (1) Ca 2+ -регулируемое включение / выключение тропонинового комплекса, (2) химические этапы в цикле кросс-мостика аденозинтрифосфатазы, (3) механика генерации силы и (4) динамика длины отдельных саркомеров. Эти исследования дают новое представление о кинетике регуляции тонких волокон и оборота поперечных мостиков, о том, как поперечные мостики преобразуют химическую энергию в механическую работу, и предполагают, что ансамбли поперечных мостиков каждого полусаркомера взаимодействуют друг с другом через полусаркомерные бордюры.Кроме того, теперь мы лучше понимаем расслабление мышц и их нарушение при определенных мышечных заболеваниях. Ключевые слова Сокращение мышц. Расслабление мышц. Сокращение миокарда. Расслабление миокарда. Миофибриллы. Саркомеры. Кальций. Регулировка тонкой нитью. Кинетика поперечного моста. Расслабление. Поперечный мост. Мышечная механика.
Силовые взаимодействия регулируются Ca 2+ через специализированные белки тонкой нити. Это является спорным, как сократительный и регуляторных систем динамически взаимодействуют, чтобы определить ход времени мышечного сокращения и расслабления.В то время как кинетику Ca 2+ -индуцированной регуляции тонких филаментов часто исследуют с использованием изолированных белков, кинетику силы обычно изучают в мышечных волокнах. Разрыв между исследованиями изолированных белков и структурированных волокон в настоящее время устраняется с помощью новейших методов, которые анализируют химическую и механическую кинетику небольших компонентов мышечного волокна, субклеточных миофибрилл, изолированных из скелетных и сердечных мышц. Сформированные из последовательно расположенных повторяющихся единиц, называемых саркомерами, миофибриллы имеют полный полностью структурированный ансамбль сократительных и регуляторных белков Ca 2+.Малый диаметр миофибрилл (несколько микрометров) облегчает анализ кинетики сокращения и расслабления саркомера, вызванных быстрыми изменениями [АТФ] или [Ca 2+]. Среди процессов, изучаемых на миофибриллах: (1) Ca 2+ -регулируемое включение / выключение тропонинового комплекса, (2) химические этапы в цикле кросс-мостика аденозинтрифосфатазы, (3) механика генерации силы и (4) динамика длины отдельных саркомеров. Эти исследования дают новое представление о кинетике регуляции тонких волокон и оборота поперечных мостиков, о том, как поперечные мостики преобразуют химическую энергию в механическую работу, и предполагают, что ансамбли поперечных мостиков каждого полусаркомера взаимодействуют друг с другом через полусаркомерные бордюры.Кроме того, теперь мы лучше понимаем расслабление мышц и их нарушение при определенных мышечных заболеваниях. Ключевые слова Сокращение мышц. Расслабление мышц. Сокращение миокарда. Расслабление миокарда. Миофибриллы. Саркомеры. Кальций. Регулировка тонкой нитью. Кинетика поперечного моста. Расслабление. Поперечный мост. Мышечная механика. Сердечный саркомер. Сердечная мышца. Сердечная функция. Кальций в клетке. Регулирование кальция. Кожаные волокна Полосатые мышцы имеют иерархически организованную архитектуру.Скелетные мышцы — это большие пучки многоядерных клеток, называемых волокнами, расположенных параллельно. Сердечная мышца состоит из сетей разветвленных одноядерных клеток, называемых миоцитами. Клетки, в свою очередь, содержат пучки миофибрилл, которые образуют сократительный аппарат. Миофибриллы расположены параллельно в волокне скелетных мышц или в разветвленных пучках внутри миоцита. На уровне отдельных миофибрилл скелетные и сердечные мышцы имеют очень похожие субструктуры. Оба типа миофибрилл состоят из наборов коротких цилиндрических повторяющихся единиц, называемых саркомерами.Саркомер — наиболее упорядоченная структура из всех клеточных органелл. Он имеет двустороннюю симметрию, то есть состоит из двух противоположных половин — полусаркомеров. Саркомер состоит из двух типов поперечно ориентированных мультибелковых каркасов, М-линии и Z-диска, которые закрепляют упорядоченную решетку из трех типов аксиальных нитей, называемых толстыми, тонкими и титиновыми нитями. Z-диски образуют внешние концы саркомера и являются общими с соседними саркомерами (рис. 1). Линия M находится в середине саркомера и разделяется двумя полусаркомерами.Середина толстых нитей закреплена на М-линии; они простираются к Z-дискам, но обычно не касаются их. Напротив, тонкие волокна прикреплены к Z-линиям, но не к M-линии. Эластичная нить тайтина также прикреплена к Z-дискам и является единственной нитью, которая образует постоянное соединение между Z-диском и M-линией; тем самым он определяет упругие свойства расслабленного саркомера [47, 86]. Каждая из тонких и толстых нитей содержит точную сборку из нескольких белков, которые вместе образуют регулируемый кальцием сократительный агрегат.Тонкие филаменты в основном состоят из актина и связанных с ним регуляторных белков, комплекса тропонина (Tn) и тропомиозина (Tm).
Сердечный саркомер. Сердечная мышца. Сердечная функция. Кальций в клетке. Регулирование кальция. Кожаные волокна Полосатые мышцы имеют иерархически организованную архитектуру.Скелетные мышцы — это большие пучки многоядерных клеток, называемых волокнами, расположенных параллельно. Сердечная мышца состоит из сетей разветвленных одноядерных клеток, называемых миоцитами. Клетки, в свою очередь, содержат пучки миофибрилл, которые образуют сократительный аппарат. Миофибриллы расположены параллельно в волокне скелетных мышц или в разветвленных пучках внутри миоцита. На уровне отдельных миофибрилл скелетные и сердечные мышцы имеют очень похожие субструктуры. Оба типа миофибрилл состоят из наборов коротких цилиндрических повторяющихся единиц, называемых саркомерами.Саркомер — наиболее упорядоченная структура из всех клеточных органелл. Он имеет двустороннюю симметрию, то есть состоит из двух противоположных половин — полусаркомеров. Саркомер состоит из двух типов поперечно ориентированных мультибелковых каркасов, М-линии и Z-диска, которые закрепляют упорядоченную решетку из трех типов аксиальных нитей, называемых толстыми, тонкими и титиновыми нитями. Z-диски образуют внешние концы саркомера и являются общими с соседними саркомерами (рис. 1). Линия M находится в середине саркомера и разделяется двумя полусаркомерами.Середина толстых нитей закреплена на М-линии; они простираются к Z-дискам, но обычно не касаются их. Напротив, тонкие волокна прикреплены к Z-линиям, но не к M-линии. Эластичная нить тайтина также прикреплена к Z-дискам и является единственной нитью, которая образует постоянное соединение между Z-диском и M-линией; тем самым он определяет упругие свойства расслабленного саркомера [47, 86]. Каждая из тонких и толстых нитей содержит точную сборку из нескольких белков, которые вместе образуют регулируемый кальцием сократительный агрегат.Тонкие филаменты в основном состоят из актина и связанных с ним регуляторных белков, комплекса тропонина (Tn) и тропомиозина (Tm).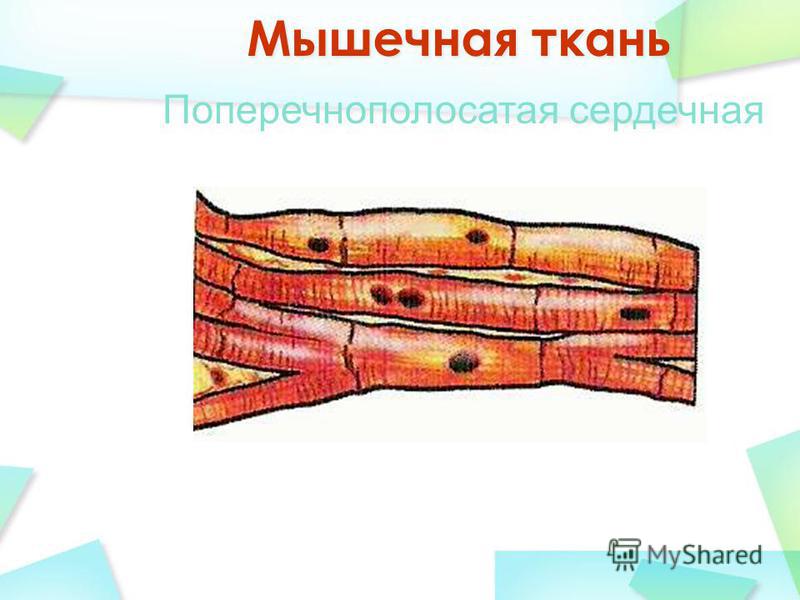 Толстые филаменты в основном состоят из миозина, легких цепей миозина и миозинсвязывающего белка C. Части миозина, называемые головками или поперечными мостиками, через равные промежутки времени выступают от основы толстой филамента к тонким филаментам. Эта структурная иерархия связывает сократительную активность от молекулярного уровня через филамент, полусаркомер, миофибриллы, клетку и до мышцы.Самая маленькая полная сократительная единица — полусаркомер одиночной миофибриллы. Полная функциональная двигательная единица — это группа мышечных волокон, управляемых одним двигательным нейроном или сердцем. На молекулярном уровне и уровне филаментов сокращение и релаксация регулируются связыванием / диссоциацией Ca 2+ с тропонином C (TnC) и от него. Связывание Ca 2+ приводит к каскаду конформационных изменений с участием TnC и других регуляторных белков тонких филаментов, тропонина I (TnI), тропонина T (TnT) и Tm; это позволяет поперечным мостикам взаимодействовать с актином [46, 75, 76].Под действием активности циклической аденозинтрифосфатазы (АТФазы) поперечные мостики воздействуют на тонкую нить, которая тянет последнюю к центру саркомера, М-линии; полусаркомер сжимается и укорачивается [59, 61, 62]. Таким образом, кинетика поперечного мостика определяет динамику укорочения конкретного полусаркомера [27]. Когда [Ca 2+] падает и Ca 2+ диссоциирует от TnC, тонкая нить инактивируется; генерирующее силу взаимодействие поперечных мостиков прекращается, и полусаркомер возвращается к своей расслабленной длине, определяемой эластичностью тайтина.Если бы ансамбль отдельных полусаркомеров действовал независимо, механизм сокращения поперечно-полосатых мышц определялся бы характеристиками одного полусаркомера. Однако кинетика цикла поперечного мостика, в частности переходы поперечных мостиков через генерирующие силу состояния, которые определяют кажущуюся скорость отрыва поперечного мостика от актина, сами по себе зависят от скорости скольжения филамента [41, 59 , 129]. Это, в свою очередь, зависит от общей силы, создаваемой всеми последовательно соединенными полусаркомерами.
Толстые филаменты в основном состоят из миозина, легких цепей миозина и миозинсвязывающего белка C. Части миозина, называемые головками или поперечными мостиками, через равные промежутки времени выступают от основы толстой филамента к тонким филаментам. Эта структурная иерархия связывает сократительную активность от молекулярного уровня через филамент, полусаркомер, миофибриллы, клетку и до мышцы.Самая маленькая полная сократительная единица — полусаркомер одиночной миофибриллы. Полная функциональная двигательная единица — это группа мышечных волокон, управляемых одним двигательным нейроном или сердцем. На молекулярном уровне и уровне филаментов сокращение и релаксация регулируются связыванием / диссоциацией Ca 2+ с тропонином C (TnC) и от него. Связывание Ca 2+ приводит к каскаду конформационных изменений с участием TnC и других регуляторных белков тонких филаментов, тропонина I (TnI), тропонина T (TnT) и Tm; это позволяет поперечным мостикам взаимодействовать с актином [46, 75, 76].Под действием активности циклической аденозинтрифосфатазы (АТФазы) поперечные мостики воздействуют на тонкую нить, которая тянет последнюю к центру саркомера, М-линии; полусаркомер сжимается и укорачивается [59, 61, 62]. Таким образом, кинетика поперечного мостика определяет динамику укорочения конкретного полусаркомера [27]. Когда [Ca 2+] падает и Ca 2+ диссоциирует от TnC, тонкая нить инактивируется; генерирующее силу взаимодействие поперечных мостиков прекращается, и полусаркомер возвращается к своей расслабленной длине, определяемой эластичностью тайтина.Если бы ансамбль отдельных полусаркомеров действовал независимо, механизм сокращения поперечно-полосатых мышц определялся бы характеристиками одного полусаркомера. Однако кинетика цикла поперечного мостика, в частности переходы поперечных мостиков через генерирующие силу состояния, которые определяют кажущуюся скорость отрыва поперечного мостика от актина, сами по себе зависят от скорости скольжения филамента [41, 59 , 129]. Это, в свою очередь, зависит от общей силы, создаваемой всеми последовательно соединенными полусаркомерами.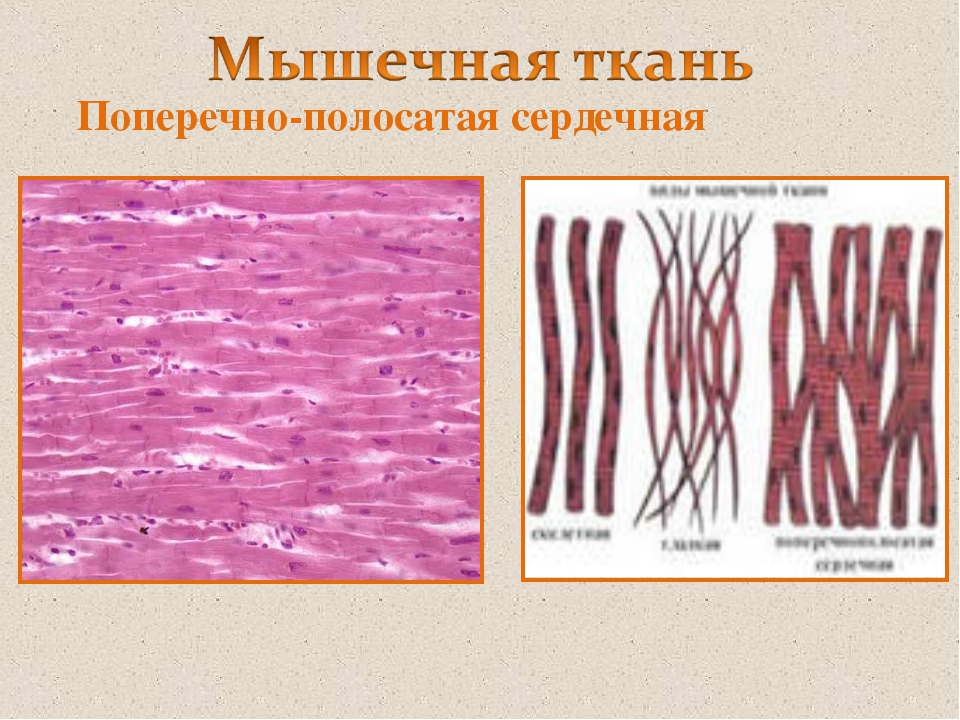 Межаркомерное соединение скольжения филаментов и отслоения поперечного моста становится очевидным, когда миофибриллы расслабляются после окончания сокращения. Быстрая релаксация происходит за счет последовательного удлинения отдельных полусаркомеров, которое пространственно распространяется вдоль миофибриллы, пока все полусаркомеры не вернут свою расслабленную длину [135, 143]. Этот последовательный процесс обеспечивает быстрое снятие механического напряжения и быструю релаксацию [117, 135, 147]. Расположение миофибрилл в волокнах и миоцитах оптимизировано для конкретной функции органа.В скелетных мышцах параллельное расположение миофибрилл и волокон передает свою силу непосредственно на сухожилия. В сердце разветвленные миоциты образуют пучки клеток с предпочтительной, но не однонаправленной ориентацией, которые спиралями обвивают полости [109]. В обеих мышцах клетки стабилизированы внеклеточным коллагеновым матриксом, который защищает их от повреждения из-за чрезмерного растяжения. Таким образом, механические характеристики мышцы являются результатом взаимодействий, происходящих на многих уровнях организации: внутримолекулярных и межмолекулярных, между толстыми и тонкими нитями, между саркомерами и между мышечными клетками и другими структурами.Поэтому сложно понять многие механизмы, лежащие в основе динамики цикла сокращения-релаксации. В принципе, кинетические параметры могут быть получены из препаратов на всех уровнях структурной иерархии, то есть от органа in vivo до изолированной молекулы. В то время как полная клеточная физиология сокращения и расслабления мышц может быть изучена только на интактных мышечных препаратах, которые включают не только саркомер, но и интактные структуры, обрабатывающие Ca 2+ [5, 67], постепенное повышение и понижение [Ca 2+] ] в этих препаратах препятствует исследованию кинетических механизмов, лежащих в основе механических характеристик мышцы.Если мы сосредоточимся на процессе генерации силы и его регуляции в саркомере, в основном есть две модели для исследования их механизмов.
Межаркомерное соединение скольжения филаментов и отслоения поперечного моста становится очевидным, когда миофибриллы расслабляются после окончания сокращения. Быстрая релаксация происходит за счет последовательного удлинения отдельных полусаркомеров, которое пространственно распространяется вдоль миофибриллы, пока все полусаркомеры не вернут свою расслабленную длину [135, 143]. Этот последовательный процесс обеспечивает быстрое снятие механического напряжения и быструю релаксацию [117, 135, 147]. Расположение миофибрилл в волокнах и миоцитах оптимизировано для конкретной функции органа.В скелетных мышцах параллельное расположение миофибрилл и волокон передает свою силу непосредственно на сухожилия. В сердце разветвленные миоциты образуют пучки клеток с предпочтительной, но не однонаправленной ориентацией, которые спиралями обвивают полости [109]. В обеих мышцах клетки стабилизированы внеклеточным коллагеновым матриксом, который защищает их от повреждения из-за чрезмерного растяжения. Таким образом, механические характеристики мышцы являются результатом взаимодействий, происходящих на многих уровнях организации: внутримолекулярных и межмолекулярных, между толстыми и тонкими нитями, между саркомерами и между мышечными клетками и другими структурами.Поэтому сложно понять многие механизмы, лежащие в основе динамики цикла сокращения-релаксации. В принципе, кинетические параметры могут быть получены из препаратов на всех уровнях структурной иерархии, то есть от органа in vivo до изолированной молекулы. В то время как полная клеточная физиология сокращения и расслабления мышц может быть изучена только на интактных мышечных препаратах, которые включают не только саркомер, но и интактные структуры, обрабатывающие Ca 2+ [5, 67], постепенное повышение и понижение [Ca 2+] ] в этих препаратах препятствует исследованию кинетических механизмов, лежащих в основе механических характеристик мышцы.Если мы сосредоточимся на процессе генерации силы и его регуляции в саркомере, в основном есть две модели для исследования их механизмов.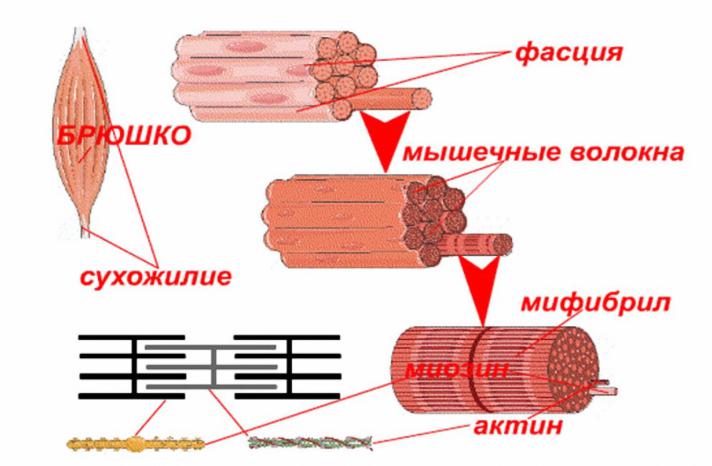 Классическая модель для изучения Са 2+ …
Классическая модель для изучения Са 2+ …
Структура скелетных мышц | SEER Training
Целая скелетная мышца считается органом мышечной системы. Каждый орган или мышца состоит из скелетной мышечной ткани, соединительной ткани, нервной ткани и крови или сосудистой ткани.
Скелетные мышцы значительно различаются по размеру, форме и расположению волокон. Они варьируются от очень крошечных прядей, таких как стремечковая мышца среднего уха, до больших масс, таких как мышцы бедра. Некоторые скелетные мышцы имеют широкую форму, а некоторые узкие. В некоторых мышцах волокна параллельны длинной оси мышцы; в некоторых они сходятся к узкой насадке; а в некоторых они косые.
Каждое волокно скелетной мышцы представляет собой отдельную цилиндрическую мышечную клетку.Отдельная скелетная мышца может состоять из сотен или даже тысяч мышечных волокон, связанных вместе и обернутых соединительно-тканевым покрытием. Каждая мышца окружена соединительнотканной оболочкой, называемой эпимизием. Фасция, соединительная ткань вне эпимизия, окружает и разделяет мышцы. Части эпимизия выступают внутрь, чтобы разделить мышцу на части. Каждый отсек содержит пучок мышечных волокон. Каждый пучок мышечных волокон называется пучком и окружен слоем соединительной ткани, называемым перимизием.Внутри пучка каждая отдельная мышечная клетка, называемая мышечным волокном, окружена соединительной тканью, называемой эндомизием.
Клетки (волокна) скелетных мышц, как и другие клетки тела, мягкие и хрупкие. Покрытие из соединительной ткани обеспечивает поддержку и защиту нежных клеток и позволяет им противостоять силам сжатия. Покрытия также обеспечивают проходы кровеносных сосудов и нервов.
Обычно эпимизий, перимизий и эндомизий выходят за пределы мясистой части мышцы, живота или желудка, образуя толстое веревочное сухожилие или широкий плоский листовой апоневроз.Сухожилие и апоневроз образуют непрямые соединения мышц с надкостницей костей или соединительной тканью других мышц. Обычно мышца охватывает сустав и прикрепляется к костям сухожилиями с обоих концов. Одна из костей остается относительно неподвижной или стабильной, в то время как другой конец движется в результате сокращения мышц.
Обычно мышца охватывает сустав и прикрепляется к костям сухожилиями с обоих концов. Одна из костей остается относительно неподвижной или стабильной, в то время как другой конец движется в результате сокращения мышц.
Скелетные мышцы имеют обильное снабжение кровеносных сосудов и нервов. Это напрямую связано с основной функцией скелетных мышц — сокращением.Прежде чем скелетное мышечное волокно сможет сократиться, оно должно получить импульс от нервной клетки. Обычно артерия и по крайней мере одна вена сопровождают каждый нерв, который проникает в эпимизий скелетной мышцы. Ветви нерва и кровеносные сосуды следуют за соединительнотканными компонентами мышцы нервной клетки и с одним или несколькими мельчайшими кровеносными сосудами, называемыми капиллярами.
Физиология человека — мышцы
Физиология человека — мышцы
Нервная система «общается» с мышцами через нервно-мышечные (также называемые мионевральными) соединениями.Эти соединения (Рисунок 1) работают
очень похоже на синапс между нейронами (видео нервно-мышечного соединения). Другими словами:
- импульс поступает на наконечник колбы,
- химический медиатор высвобождается из пузырьков (каждая из которых содержит 5000-10 000 молекул ацетилхолина) и распространяется через нервно-мышечные
расщелина, - молекулы передатчика заполняют рецепторные участки в мембране мышцы и увеличивают проницаемость мембраны для натрия,
- натрия диффундирует внутрь и мембранный потенциал становится менее отрицательным,
- и, если пороговый потенциал достигнут, возникает потенциал действия,
импульс проходит по мембране мышечной клетки, и мышца сокращается.
Затем
Некоторые мышцы (скелетные) не будут сокращаться без стимуляции.
нейронами; другие мышцы (гладкие и сердечные) будут сокращаться без
нервное возбуждение, но на их сокращение может влиять нервная
система. Таким образом, нервная и мышечная системы тесно взаимосвязаны.
Давайте теперь сосредоточимся на мышцах — какова их структура и как она работает.
Изображение нервно-мышечного соединения в большом увеличении (Hirsch 2007).
Характеристики мышцы:
- возбудимость — реагирует на раздражители (например, нервные импульсы)
- сократимость — возможность укорочения
- растяжимость — растягивается при вытягивании
- эластичность — имеет тенденцию возвращаться к исходной форме и длине после сжатия или растяжения
Функции мышцы:
- движение
- поддержание осанки
- производство тепла
Типы мышц:
- каркас:
- прикреплен к костям и перемещает скелет
- также называется поперечно-полосатой мышцей (из-за его внешнего вида под микроскопом, как показано на фото слева)
- произвольная мышца
- гладкая (фото справа)
- Непроизвольная мышца
- мышца внутренних органов (e.g., в стенках кровеносных сосудов, кишечника и других «полых» структур и органов тела)
- сердечный:
- Сердечная мышца
- принудительное
Строение скелетных мышц
Структура скелетных мышц:
Скелетные мышцы обычно прикрепляются к кости сухожилиями, состоящими из соединительной ткани.Эта соединительная ткань также покрывает всю мышцу и называется эпимизием.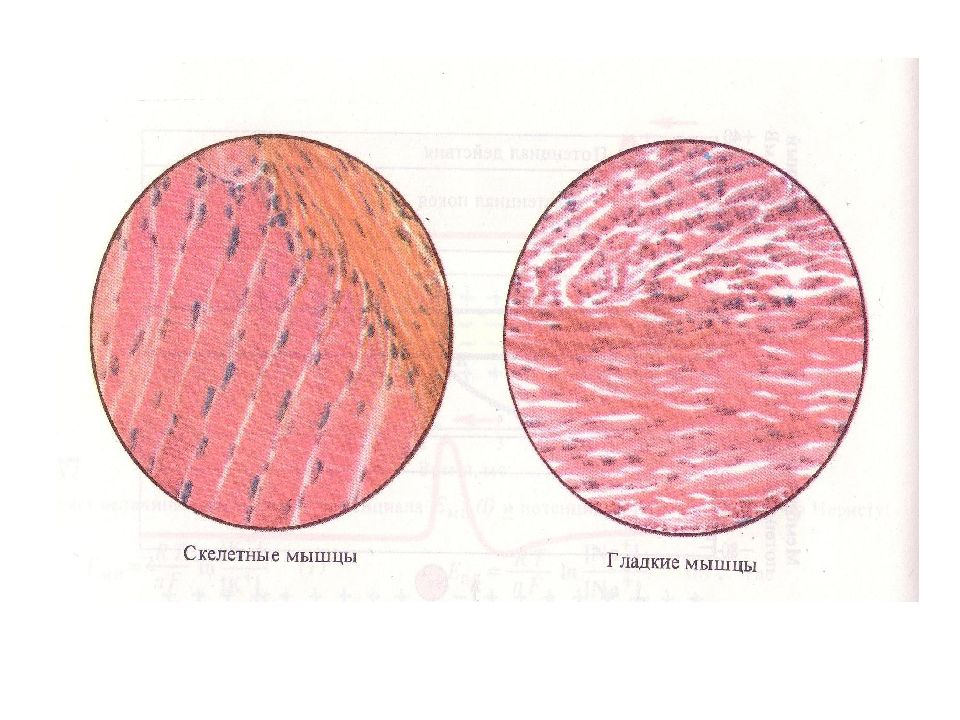 Скелетные мышцы состоят из множества субъединиц или пучков, называемых фасиклами (или пучками). Фасцикулы также окружены соединительной тканью (называемой перимизием), и каждый пучок состоит из многочисленных мышечных волокон (или мышечных клеток). Мышечные клетки, окруженные эндомизием, состоят из множества фибрилл (или миофибрилл), а эти миофибриллы состоят из длинных белковых молекул, называемых миофиламентами. В миофибриллах есть два типа миофиламентов: толстые миофиламенты и тонкие миофиламенты.
Скелетные мышцы состоят из множества субъединиц или пучков, называемых фасиклами (или пучками). Фасцикулы также окружены соединительной тканью (называемой перимизием), и каждый пучок состоит из многочисленных мышечных волокон (или мышечных клеток). Мышечные клетки, окруженные эндомизием, состоят из множества фибрилл (или миофибрилл), а эти миофибриллы состоят из длинных белковых молекул, называемых миофиламентами. В миофибриллах есть два типа миофиламентов: толстые миофиламенты и тонкие миофиламенты.
Источник: Википедия.
Скелетные мышцы значительно различаются по размеру, форме и расположению волокон. Они варьируются от очень крошечных прядей, таких как стремечковая мышца среднего уха, до больших масс, таких как мышцы бедра. Скелетные мышцы могут состоять из сотен или даже тысяч мышечных волокон, связанных вместе и завернутых в соединительнотканный покров. Каждая мышца окружена соединительнотканной оболочкой, называемой эпимизием.Фасция, соединительная ткань вне эпимизия, окружает и разделяет мышцы. Части эпимизия выступают внутрь, чтобы разделить мышцу на части. Каждый отсек содержит пучок мышечных волокон. Каждый пучок мышечных волокон называется пучком и окружен слоем соединительной ткани, называемым перимизием. Внутри пучка каждая отдельная мышечная клетка, называемая мышечным волокном, окружена соединительной тканью, называемой эндомизием. Скелетные мышцы имеют обильное снабжение кровеносных сосудов и нервов.Прежде чем скелетное мышечное волокно сможет сократиться, оно должно получить импульс от нейрона. Обычно артерия и по крайней мере одна вена сопровождают каждый нерв, который проникает в эпимизий скелетной мышцы. Ветви нерва и кровеносные сосуды следуют за соединительнотканными компонентами мышцы нервной клетки и с одним или несколькими мельчайшими кровеносными сосудами, называемыми капиллярами (Источник: training.seer.cancer.gov). |
Клеточная мембрана мышечной клетки называется сарколеммой, и эта мембрана, как и мембрана нейронов, поддерживает мембранный потенциал. Итак, импульсы проходят по мембранам мышечных клеток так же, как и по мембранам нервных клеток. Однако «функция» импульсов в мышечных клетках — вызывать сокращение. Чтобы понять, как сокращается мышца, вам нужно немного узнать о структуре мышечных клеток.
Итак, импульсы проходят по мембранам мышечных клеток так же, как и по мембранам нервных клеток. Однако «функция» импульсов в мышечных клетках — вызывать сокращение. Чтобы понять, как сокращается мышца, вам нужно немного узнать о структуре мышечных клеток.
Скелетная мышца — это мышца, прикрепленная к скелету. Сотни или тысячи мышечных волокон (клеток) связываются вместе, образуя отдельные скелетные мышцы.Мышечные клетки представляют собой длинные цилиндрические структуры, которые связаны плазматической мембраной (сарколеммой) и вышележащей базальной пластинкой, и когда они сгруппированы в пучки (пучки), они составляют мышцы. Сарколемма образует физический барьер против внешней среды, а также передает сигналы между внешней средой и мышечной клеткой. Саркоплазма — это специализированная цитоплазма мышечной клетки, которая содержит обычные субклеточные элементы наряду с аппаратом Гольджи, множеством миофибрилл, модифицированным эндоплазматическим ретикулумом, известным как саркоплазматический ретикулум (SR), миоглобином и митохондриями.Поперечные (Т) -рубочки инвагинируют сарколемму, позволяя импульсам проникать в клетку и активировать SR. Как показано на рисунке, SR образует сеть вокруг миофибрилл, накапливая и обеспечивая Ca 2+ , который необходим для сокращения мышц. Миофибриллы — это сократительные единицы, которые состоят из упорядоченного расположения продольных миофиламентов. Миофиламенты могут быть толстыми (состоящими из миозина) или тонкими (состоящими в основном из актина).Характерные «полосы» скелетных и сердечных мышц легко наблюдаются при световой микроскопии в виде чередующихся светлых и темных полос на продольных срезах. Светлая полоса (известная как I-полоса) состоит из тонких нитей, тогда как темная полоса (известная как A-полоса) состоит из толстых нитей. Z-линия (также известная как Z-диск или Z-полоса) определяет боковую границу каждой саркомерной единицы. |
 Сокращение саркомера происходит, когда Z-линии сближаются, заставляя миофибриллы сокращаться, и, следовательно, сокращается вся мышечная клетка, а затем и вся мышца (Источник: Davies and Nowak 2006).
Сокращение саркомера происходит, когда Z-линии сближаются, заставляя миофибриллы сокращаться, и, следовательно, сокращается вся мышечная клетка, а затем и вся мышца (Источник: Davies and Nowak 2006).SARCOLEMMA имеет уникальную особенность: в ней есть дыры. Эти «отверстия» ведут в трубки, называемые ПОПЕРЕЧНЫМИ ТРУБКАМИ (или сокращенно Т-ТРУБКАМИ). Эти канальцы проходят вниз в мышечную клетку и огибают МИОФИБРИЛЫ. Однако эти канальцы НЕ открываются внутрь мышечной клетки; они полностью проходят и открываются где-то еще на сарколемме (т. е. эти канальцы не используются, чтобы вводить и выводить предметы в мышечную клетку).Функция T-TUBULES — проводить импульсы с поверхности клетки (SARCOLEMMA) вниз в клетку и, в частности, в другую структуру клетки, называемую SARCOPLASMIC RETICULUM.
Мышечное волокно возбуждается через двигательный нерв, который генерирует потенциал действия, который распространяется по поверхностной мембране (сарколемме) и поперечной трубчатой системе в более глубокие части мышечного волокна. Белок рецептора (DHP) воспринимает деполяризацию мембраны, изменяет ее конформацию и активирует рецептор рианодина (RyR), который высвобождает Ca 2+ из SR.Затем Ca 2+ связывается с тропонином и активирует процесс сокращения (Jurkat-Rott and Lehmann-Horn 2005).
Мембраны саркоплазматического ретикулума (SR) в непосредственной близости от Т-канальца. «RyR» — это белки, способствующие высвобождению кальция из SR, «SERCA2» — это белки, которые помогают транспортировать кальций в SR (Brette and Orchard 2007).
SARCOPLASMIC RETICULUM (SR) немного похож на эндоплазматический ретикулум других клеток, например.г., он полый. Но основная функция SARCOPLASMIC RETICULUM — ХРАНЕНИЕ ИОНОВ КАЛЬЦИЯ. Саркоплазматический ретикулум очень богат клетками скелетных мышц и тесно связан с МИОФИБРИЛАМИ (и, следовательно, МИОФИЛАМЕНТАМИ). Мембрана SR хорошо приспособлена для обработки кальция: существуют «насосы» (активный транспорт) для кальция, так что кальций постоянно «закачивается» в SR из цитоплазмы мышечной клетки (так называемая SARCOPLASM). В результате в расслабленной мышце наблюдается очень высокая концентрация кальция в SR и очень низкая концентрация в саркоплазме (и, следовательно, среди миофибрилл и миофиламентов).Кроме того, в мембране есть специальные отверстия или «ворота» для кальция. В расслабленной мышце эти ворота закрыты, и кальций не может проходить через мембрану. Итак, кальций остается в SR. Однако, если импульс проходит по мембране SR, кальциевые «ворота» открываются, и, следовательно, кальций быстро диффундирует из SR в саркоплазму, где расположены миофибриллы и миофиламенты. Как вы увидите, это ключевой шаг в сокращении мышц.
Миофибриллы состоят из миофиламентов двух типов: толстых и тонких.В скелетных мышцах эти миофиламенты расположены очень правильным и точным образом: толстые миофиламенты обычно окружены 6 тонкими миофиламентами (вид с торца). На виде сбоку тонкие миофиламенты можно увидеть над и под каждым толстым миофиламентом.
Поперечный разрез миофибрилл, показывающий расположение толстых и тонких миофиламентов.
Бар = 100 нм. Изображение Widrick et al. (2001)
Источник: Цховребова, Триник (2003).
Строение мышц
Каждая миофибрилла состоит из множества субъединиц, выстроенных встык. Эти
субъединицы, конечно, состоят из миофиламентов и называются SARCOMERES.
Рисунки выше и ниже показывают только очень маленькую часть всего
длина миофибриллы, поэтому вы можете увидеть только один полный SARCOMERE.
В каждом саркомере тонкие миофиламенты проходят с каждого конца.Толстый
миофиламенты находятся в середине саркомера и не расширяются
до конца. Из-за такого расположения при просмотре скелетных мышц
под микроскопом концы саркомера (там, где только тонкие миофиламенты
найдены) кажутся светлее, чем центральная часть (которая темная, потому что
наличия толстых миофиламентов). Таким образом, миофибрилла имеет чередующиеся
светлые и темные области, потому что каждая состоит из множества выстроенных в линию саркомеров
концы с концами. Вот почему скелетная мышца называется ПОЛОСКОЙ МЫШЦЫ (т.е.,
чередующиеся светлые и темные участки выглядят как полосы или полосы).
Светлые области называются I-BAND, а более темные — A-BAND.
Рядом с центром каждой I-BAND есть тонкая темная линия, называемая Z-LINE (или
Z-мембрана на рисунке ниже). Z-LINE — это место, где соседние саркомеры
сходятся вместе, и тонкие миофиламенты соседних саркомеров перекрываются
немного. Таким образом, саркомер можно определить как область между Z-линиями.
Используется с разрешения Джона
W.Кимбалл
Толстые миофиламенты состоят из белка под названием МИОЗИН. Каждый МИОЗИН
молекула имеет хвост, который образует ядро толстой миофиламента плюс
головка, выступающая из сердцевины нити. Эти головы MYOSIN
также обычно называют ПЕРЕКРЕСТНЫМИ МОСТАМИ.
МИОЗИН
HEAD имеет несколько важных характеристик:
- у него есть АТФ-связывающие сайты, в которые помещаются молекулы АТФ.АТФ представляет собой
потенциальная энергия. - у него есть сайты связывания ACTIN, в которые помещаются молекулы ACTIN. Актин
является частью тонкой миофиламента и будет обсуждаться более подробно в ближайшее время. - он имеет «шарнир» в том месте, где он выходит из ядра толстой миофиламента.
Это позволяет голове поворачиваться вперед и назад, а «поворот»
как будет описано вкратце, что на самом деле вызывает сокращение мышц.
Тонкие миофиламенты состоят из 3 типов белков: АКТИН, ТРОПОНИН,
и ТРОПОМИОСИН.
Молекулы актина (или G-актин, как указано выше) имеют сферическую форму и образуют длинные
цепи. Каждая тонкая миофиламент содержит две такие цепочки, которые обвиваются вокруг
друг с другом. Молекулы ТРОПОМИОЗИНА — это одиночные, тонкие молекулы, которые обвивают
цепочка ACTIN. В конце каждого тропомиозина находится молекула ТРОПОНИНА.
Молекулы ТРОПОМИОЗИНА и ТРОПОНИНА связаны друг с другом.Каждый
из этих 3 белков играет ключевую роль в сокращении мышц:
- АКТИН — когда актин объединяется с ГОЛОВКОЙ МИОЗИНА, АТФ, связанный с
голова распадается на АДФ. Эта реакция высвобождает энергию, которая вызывает
МИОЗИН ГОЛОВА ПОВОРОТА.
- ТРОПОМИОЗИН — В расслабленной мышце МИОЗИНОВЫЕ ГОЛОВКИ толстой миофиламента
лежат против молекул ТРОПОМИОЗИНА тонкой миофиламента.Пока
ГОЛОВКИ МИОЗИНА остаются в контакте с ТРОПОМИОЗИНОМ, ничего не происходит (т. Е.
мышца остается расслабленной).
- ТРОПОНИН — молекулы тропонина имеют участки связывания для ионов кальция. Когда
ион кальция заполняет это место, он вызывает изменение формы и положения
ТРОПОНИНА. И когда ТРОПОНИН смещается, он притягивает ТРОПОМИОЗИН, к которому
он прилагается. Когда ТРОПОМИОЗИН перемещается, ГОЛОВА МИОЗИНА, которая касалась
теперь тропомиозин вступает в контакт с лежащей в основе молекулой ACTIN.
Нити раздвижные
Сокращение мышц
1 — Поскольку скелетная мышца является произвольной мышцей, для ее сокращения требуется
нервный импульс. Итак, шаг 1 в сокращении — это когда импульс передается.
от нейрона к САРКОЛЕММЕ мышечной клетки.
2 — Импульс проходит по САРКОЛЕММЕ и вниз
ТРУБКИ.От Т-ТРУБОК импульс переходит в САРКОПЛАЗМИЧЕСКУЮ
РЕТИКУЛУМ.
3 — Когда импульс движется по саркоплазматической сети (SR),
кальциевые ворота в мембране СР открываются. В результате КАЛЬЦИЙ диффундирует
вне SR и среди миофиламентов.
4 — Кальций
заполняет сайты связывания в молекулах ТРОПОНИНА. Как отмечалось ранее,
это
изменяет форму и положение ТРОПОНИНА, что, в свою очередь, вызывает
движение присоединенной молекулы ТРОПОМИОЗИНА.
5 — Движение ТРОПОМИОЗИНА позволяет ГОЛОВЕ МИОЗИНА контактировать с АКТИНОМ (анимации: сокращение миофиламентов и разрушение АТФ и движение поперечного моста).
6 — При контакте с ACTIN ГОЛОВКА МИОЗИНА поворачивается.
7 — Во время поворота ГОЛОВКА МИОЗИНА надежно прикреплена к ACTIN.
Итак, когда ГОЛОВКА поворачивается, она тянет ACTIN (и, следовательно, весь
тонкая миофиламент) вперед. (Очевидно, одна ГОЛОВА МИОЗИНА не может тянуть
вся тонкая миофиламент.Многие МИОЗИНОВЫЕ ГОЛОВКИ вращаются одновременно,
или почти так, и их коллективных усилий достаточно, чтобы вытащить все
тонкий миофиламент).
8 — В конце вертлюга АТФ входит в сайт связывания на
поперечный мост и это разрывает связь между поперечным мостом (миозин)
и актин. ГОЛОВА МИОЗИНА затем поворачивается назад. Когда он поворачивается назад, ATP
распадается на ADP & P, и поперечный мостик снова связывается с актином.
молекула.
9 — В результате HEAD снова прочно привязан к ACTIN.
Однако, поскольку ГОЛОВА не была прикреплена к актину, когда она поворачивалась назад,
HEAD будет связываться с другой молекулой ACTIN (т.е.
на тонком миофиламенте). После присоединения HEAD к ACTIN перекрестный мост
снова вертится, ТАК ШАГ
7 ПОВТОРЯЕТСЯ.
Пока присутствует кальций (присоединенный к ТРОПОНИНУ), шаги с 7 по
9 продолжу. И при этом тонкая миофиламент «тянется»
МИОЗИНОВЫМИ ГОЛОВКАМИ толстой миофиламента.Таким образом, THICK & THIN
миофиламенты действительно скользят
мимо друг друга. При этом расстояние между Z-линиями
саркомер уменьшается. По мере того как саркомеры становятся короче, миофибриллы, конечно,
становится короче. И, очевидно, мышечные волокна (и вся мышца) становятся
короче.
Скелетные мышцы расслабляются, когда прекращается нервный импульс. Нет импульса значит
что мембрана SARCOPLASMIC RETICULUM больше не проницаема
к кальцию (т.д., отсутствие импульса означает, что КАЛЬЦИЕВЫЕ ВОРОТА закрываются). Итак, кальций
больше не распространяется. НАСОС КАЛЬЦИЯ в мембране теперь будет транспортировать
кальций обратно в SR. Когда это происходит, ионы кальция покидают связывающую
сайты, посвященные МОЛЕКУЛАМ ТОПОНИНА. Без кальция ТРОПОНИН возвращается к своему
исходная форма и положение, как и прилагаемый ТРОПОМИОЗИН. Это означает
что ТРОПОМИОЗИН теперь снова на месте, в контакте с ГОЛОВКОЙ МИОЗИНА.
Таким образом, головка MYOSIN больше не контактирует с ACTIN и, следовательно,
мышца перестает сокращаться (т.э., расслабляет).
Сокращение
Итак, в большинстве случаев кальций является «переключателем», который заставляет мышцы
«включение и выключение» (сокращение и расслабление). Когда мышца используется для расширенного
Период, запасы АТФ могут уменьшиться. Когда концентрация АТФ в мышцах снижается,
ГОЛОВКИ МИОЗИНА остаются связанными с актином и больше не могут поворачиваться. Это снижение
в уровне АТФ в мышцах вызывает МЫШЕЧНУЮ УСТАЛОСТЬ. Хотя кальций
все еще присутствует (и нервный импульс передается мышце),
сокращение (или, по крайней мере, сильное сокращение) невозможно.
Анимация, иллюстрирующая сокращение мышц:
Сокращение мышечных клеток
Сокращение скелетных мышц
Механизм скольжения нити при сокращении миофибриллы (Wiley)
Потенциалы действия и сокращение мышц
Виды сокращений:
- 1 — изотонический — напряжение или сила, создаваемая мышцей, больше
чем нагрузка и мышца укорачиваются
2 — изометрический — нагрузка больше, чем создаваемое натяжение или сила
мышцами и мышца не укорачивается
Twitch
— ответ скелетной мышцы на однократную стимуляцию (или действие
потенциал):
- латентный период — без изменения длины; время, в течение которого движется импульс
вдоль сарколеммы и вниз по Т-канальцам до саркоплазматической сети, кальций
высвобождается и так далее (другими словами, мышцы не могут сокращаться мгновенно!) - период схватывания — растяжение увеличивается (поперечные перемычки качаются)
- период релаксации — мышцы расслабляются (напряжение уменьшается) и имеет тенденцию к возвращению
до исходной длины
Важной характеристикой скелетных мышц является их способность сокращаться.
в разной степени.Мышца, как и бицепс, сокращается с разной степенью
силы в зависимости от обстоятельств (это также называется
отклик). Мышцы делают это с помощью процесса, называемого суммированием, а именно:
мотором
единичное суммирование и волновое суммирование.
Motor Unit Sumpting — степень сокращения скелета
На мышцу влияет количество стимулируемых двигательных единиц (с
двигательная единица, представляющая собой мотонейрон, плюс все мышечные волокна, которые он иннервирует;
см. диаграмму ниже).Скелетные мышцы состоят из множества двигательных единиц и,
следовательно, стимуляция большего количества двигательных единиц вызывает более сильное сокращение.
Волновое суммирование — увеличение частоты, с которой мышца
стимулируется, увеличивает силу сокращения. Это проиллюстрировано
в (б). При быстрой стимуляции (настолько быстрой, что мышца не полностью
расслабиться между последовательными стимуляциями), мышечное волокно повторно стимулируется
пока еще есть некоторая сократительная активность.В результате получается
«суммирование» сократительной силы. Кроме того, при быстрой стимуляции
между последовательными стимуляциями не хватает времени, чтобы удалить все
кальций из саркоплазмы. Итак, с помощью нескольких последовательных стимуляций,
уровень кальция в саркоплазме повышается. Больше кальция — значит активнее
поперечные мосты и, следовательно, более сильное сжатие. (Wiley анимация)
Если мышечное волокно стимулируется так быстро, что оно не расслабляется при
между раздражителями происходит плавное устойчивое сокращение, называемое столбняк (проиллюстрировано прямой линией в c выше и на диаграмме
ниже).
Используется с разрешения Джона
В. Кимбалл
ГЛАДКАЯ МЫШЦА:
- непроизвольная мышца; иннервируется вегетативной нервной системой (висцеральной
эфферентные волокна) - в основном обнаруживается в стенках полых органов и трубок
- Веретенообразные ячейки обычно размещаются в листах
- не имеют Т-канальцев и имеют очень маленький саркоплазматический ретикулум
- не содержат саркомеров (поэтому не имеют бороздок), но состоят из
толстые и тонкие миофиламенты.Тонкие нити в гладкой мускулатуре не содержат
тропонин. - не связывается с тропонином, а, скорее, с белком под названием кальмодулин.
Комплекс кальций-кальмодулин «активирует» миозин, который затем связывается с актином, и начинается сокращение (поворот поперечных мостиков).
Клетки
Клетки
Кальций
Два типа гладких мышц:
1 — висцеральный,
или унитарные, гладкие мышцы
- найдены в стенках полых органов (напр.г., мелкие кровеносные сосуды, пищеварительные
тракта, мочевыводящей системы и репродуктивной системы) - несколько волокон сжимаются как единое целое (потому что импульсы легко проходят через
щелевые переходы от клетки к клетке) и, в некоторых случаях, самовозбудимые
(генерируют спонтанные потенциалы действия и сокращения)
- 2 — многокомпонентная гладкая мышца
- состоит из двигательных единиц, которые активируются нервной стимуляцией
- обнаружен в стенках крупных кровеносных сосудов, в глазу (корректирует форму
линзы, чтобы разрешить размещение и размер зрачка, чтобы отрегулировать
количество света, попадающего в глаз) и у основания волосяного фолликула
(мышцы «гусиная шишка»)
Полезные ссылки:
Актин
Миозиновая анимация
Введение
к физиологии мышц и дизайну
Макгроу-Хилл:
Скелетно-мышечная система
Цитированная литература:
Бретт, Ф., и К. Орчард. 2007. Возрождение исследования сердечных Т-канальцев. Физиология 22: 167-173.
Дэвис, К. Э., и К. Дж. Новак. 2006. Молекулярные механизмы мышечных дистрофий: старые и новые игроки. Nature Reviews Molecular Cell Biology 7: 762-773.
Хирш, Н. П. 2007. Нервно-мышечные соединения в здоровье и болезни. Британский журнал анестезии 99: 132-138.
Хопкинс, П. М. 2005. Основы анестезии, второе изд. Мосби, Лондон.
Юркат-Ротт, К., и Ф. Леманн-Хорн. 2005. Мышечные каналопатии и критические точки в функциональных и геномных исследованиях. Журнал клинических исследований 115: 2000-2009.
Цховребова Л. и Дж. Триник. 2003. Титин: собственность и семейные отношения. Nature Reviews Molecular Cell Biology 4: 679-689.
Видрик, Дж. Дж., Дж. Дж. Роматовски, К. М. Норенберг, С. Т. Кнут, Дж. Л. В. Бейн, Д. А. Райли, С. В. Траппе, Т. А. Траппе, Д. Л. Костилл и Р. Х. Фиттс. 2001. Функциональные свойства медленных и быстрых волокон икроножной мышцы после 17-дневного космического полета.Журнал прикладной физиологии 90: 2203-2211.
Назад
к программе BIO 301
Лекция
Примечания 4 — Защита крови и тела
Различение трех типов мышечной ткани
Информация
Мышечная ткань — третья из четырех основных категорий тканей животных. Мышечная ткань подразделяется на три большие категории: скелетных, мышц, сердечных, мышц и гладких, мышц.Три типа мышц можно различить как по их расположению, так и по микроскопическим характеристикам.
Скелетная мышца прикреплена к костям. Он состоит из длинных многоядерных волокон . Волокна проходят по всей длине мышцы, из которой они происходят, и поэтому обычно слишком длинные, чтобы их концы были видны под микроскопом. Волокна относительно широкие и очень длинные, но неразветвленные, . Волокна не являются отдельными клетками, а образуются в результате слияния тысяч клеток-предшественников.Вот почему они такие длинные и почему отдельные волокна многоядерные (одно волокно имеет много ядер). Ядра обычно упираются в край волокна. В скелетных мышцах имеется штрихов и полосок. Это чередующиеся темные и светлые полосы, перпендикулярные краю волокна, которые присутствуют по всей длине волокна.
Сердечная мышца находится только в сердце. Его волокна длиннее, чем ширина, и имеют поперечнополосатую форму , , как волокна скелетных мышц.Но, в отличие от волокон скелетных мышц, волокна сердечной мышцы имеют отдельные концы, называемые интеркалированными дисками . Это темные линии, идущие от одной стороны волокна к другой. Вставные диски не намного толще полосок, но по этой причине они обычно темнее и так отчетливы. Одно волокно сердечной мышцы — это материал между двумя вставленными дисками. Волокна сердечной мышцы состоят из одноядерных, , только с одним ядром на волокно, и иногда они могут быть разветвленными, .
Гладкая мускулатура встречается в стенках внутренних органов, таких как органы пищеварительного тракта, кровеносные сосуды и другие. Он состоит из мононуклеарных волокон с коническими краями . Под микроскопом на гладкой мускулатуре полосок не видно. Поскольку гладкие мышцы часто оборачиваются вокруг органа, с которым они связаны, бывает трудно найти целое гладкомышечное волокно в профиле в срезе ткани на предметном стекле микроскопа. Большинство волокон будет разрезано под углом или будет трудно попасть в одну плоскость фокуса, но небольшой поиск обычно может выявить некоторые из них, при этом все определяющие характеристики будут видны.
Лаборатория 5 Упражнения 5,6
- На каждой из трех микрофотографий ниже укажите, какой тип мышцы присутствует. Перечислите определяющие визуальные характеристики этого типа мышц и нарисуйте стрелки к особенностям на фотографии, которые иллюстрируют каждую характеристику.
Анатомия мышечных клеток — Схема анатомического рисунка
Чтобы активировать мышцу, мозг посылает импульс по нерву.Мышечная клетка, технически известная как миоцит.
Анатомия гладкомышечных клеток расслабленной и сокращенной гладкой
Профессор Дэйв объясняет 89683 просмотра.
Анатомия мышечных клеток . Определение мышечных клеток мышечных клеток. Мышечные клетки редко действуют в одиночку. Мышечные органы работают по принципу дифференцированной силы моторных единиц. Функциональная единица моторной единицы мышечной системы, отдельный мотонейрон и все мышечные клетки, которые он иннервирует. может иннервировать от нескольких до сотен мышц.Из-за его нитевидной формы.
Анатомический тест 5152 62 семестра. В организме человека есть 3 типа мышечных клеток. Скелетная мышца, сердечная мышца и гладкая мышца.
Функция мышечной клетки. Как видно на изображении ниже, мышечная клетка представляет собой компактный пучок. Он имеет форму волокна, потому что он длиннее, чем ширина.
Строение мышечной клетки. Расположение белковых волокон внутри клеток вызывает появление этих светлых и темных полос.Это ядра.
Определенные пороки сердца. Миокард сердечной мышцы. Анатомия и физиология чаи руководство по приготовлению чая.
Почему отдельная клетка скелетных мышц называется мышечным волокном. Скелетная мышца или произвольная мышца крепится сухожилиями или апоневрозами в нескольких местах. Какие качества волокна скелетных мышц могут меняться с возрастом.
Штрихи указывают на то, что мышечная клетка очень сильна в отличие от висцеральных мышц.Гладкая или непроизвольная мышца находится в стенках таких органов и структур, как. У позвоночных выделяют три типа мышечной ткани.
Внутри мышечной ткани есть три отдельные группы тканей. Клетки скелетных мышц — длинные цилиндрические, многоядерные и поперечно-полосатые. Мембрана мышечной клетки — это сарколемма.
Каждая из этих групп тканей состоит из специализированных клеток, которые придают ткани ее уникальные свойства.Потенциал действия саркомеров и продолжительность нервно-мышечного соединения. Мышечная ткань — один из четырех основных типов тканей, который играет жизненно важную роль в обеспечении движения и выработки тепла в органах тела.
Клетки сердечной мышечной ткани имеют поперечно-полосатые, то есть они кажутся светлыми и темными полосами при просмотре под световым микроскопом. Каждое ядро регулирует обмен веществ. И снова этот эндомизий, как и перимизий, содержит нервы и кровеносные сосуды, которые могут помочь проводить нейронные сигналы и кровь к отдельным миофибриллам и соединительной ткани, которая находится здесь.
Клетки скелетных мышц Мышечные клетки, обычно известные как миоциты, представляют собой клетки, составляющие мышечную ткань. Механизм сокращения мышц. А затем, если вы возьмете поперечное сечение, внутри есть трубки, которые называются миофибриллами.
Таким образом, мы можем назвать это мышечной клеткой, но мы также можем назвать это миомио, что означает мышечное волокно.
Анатомия
Физиология анатомии человека в Free Ed Net
2 Нервные и мышечные клетки
Физиология скелетных мышц
Упражнение 14 Микроскопическая анатомия и организация скелета
Методика создания мышечных клеток человека
Интерактивное руководство по анатомии
Мышечные клетки в лаборатории
Внутренние части мышечной ткани Складская иллюстрация
Основы мышц Характеристики и компоненты
Анатомия скелетных мышц Волокно Видео Академия Хана
10 2 Анатомия и физиология скелетных мышц
Гладкая мышечная анатомия и анатомия мышц
Структура мышцы под наукой микроскопа
Диаграмма
мышечных волокон M uscle Fiber Cell Myofibril
Анатомия мышечной клетки
Скелетная мышечная клетка Стоковые Фото Скелетная мышечная клетка
Хордатная анатомия Хордатная анатомия Сравнительная мышца
PPT Мышечная система Скелетная мышца Ткань и мышечная ткань
Видеоанатомия скелетной мускулатуры
Мышечная анатомия
анатомии мышечной клетки Стоковые Изображения Фотография
физиологии мышечной биологии Фундаментальная наука Orthobullets
Ch 09 Общая терминология мышц
Маркированная диаграмма поперечно-полосатой мышцы говяжьей мышцы
19 2 анатомии сердечной мышцы и электрической активности и
типов Характеристики мышечных клеток Расположение Роли Kenhub
Анатомия мышечных клеток Физиология Обзор Контрольный лист 90 003
Анатомия мышечных клеток Физиология Контрольный лист опроса
4.4 мышечной ткани — анатомия и физиология
Цели обучения
Опишите характеристики мышечной ткани и то, как они определяют функцию мышц.
К концу этого раздела вы сможете:
- Определите три типа мышечной ткани
- Сравните и сопоставьте функции каждого типа мышечной ткани
Мышечная ткань обладает свойствами, позволяющими двигаться. Мышечные клетки возбудимы; они реагируют на раздражитель.Они сокращаются, то есть могут укорачиваться и создавать тянущее усилие. При прикреплении между двумя подвижными объектами, такими как две кости, сокращение мышц заставляет кости двигаться. Некоторые движения мышц являются произвольными, что означает, что они находятся под сознательным контролем. Например, человек решает открыть книгу и прочитать главу по анатомии. Другие движения являются непроизвольными, то есть они не находятся под сознательным контролем, например, сужение зрачка при ярком свете. Мышечная ткань подразделяется на три типа в зависимости от структуры и функции: скелетная, сердечная и гладкая (Таблица 4.2).
| Тип мышц | Конструкционные элементы | Функция | Расположение |
| Скелет | Длинное цилиндрическое волокно, полосатое, с множеством периферических ядер | Произвольное движение, производит тепло, защищает органы | Прикрепляется к костям и вокруг участков входа и выхода тела (например, рта, ануса) |
| Сердечный | Короткое, разветвленное, поперечно-полосатое, одно центральное ядро | Контракты на перекачку крови | Сердце |
| Гладкая | Короткое, веретенообразное, без явной штриховки, по одному ядру в каждом волокне | Непроизвольное движение, перемещение пищи, непроизвольный контроль дыхания, перемещение выделений, регулирование кровотока в артериях путем сокращения | Стенки основных органов и проходов |
Скелетная мышца прикреплена к костям, и ее сокращение делает возможным передвижение, мимику, осанку и другие произвольные движения тела.Сорок процентов вашей массы тела составляют скелетные мышцы. Скелетные мышцы выделяют тепло как побочный продукт своего сокращения и, таким образом, участвуют в тепловом гомеостазе. Дрожь — это непроизвольное сокращение скелетных мышц в ответ на более низкую, чем обычно, температуру тела. Мышечная клетка или миоцит развивается из миобластов, происходящих из мезодермы. Миоциты и их количество остаются относительно постоянными на протяжении всей жизни. Ткань скелетных мышц состоит из пучков, окруженных соединительной тканью.Под световым микроскопом мышечные клетки кажутся полосатыми с множеством ядер, сдавленных вдоль мембран. Стройность возникает из-за регулярного чередования сократительных белков актина и миозина, а также структурных белков, которые связывают сократительные белки с соединительными тканями. Клетки являются многоядерными в результате слияния множества миобластов, которые сливаются, образуя каждое длинное мышечное волокно.
Сердечная мышца образует сократительные стенки сердца. Клетки сердечной мышцы, известные как кардиомиоциты, также кажутся полосатыми под микроскопом.В отличие от волокон скелетных мышц кардиомиоциты представляют собой одиночные клетки с одним центрально расположенным ядром. Основная характеристика кардиомиоцитов состоит в том, что они сокращаются в соответствии со своим собственным ритмом без внешней стимуляции. Кардиомиоциты прикрепляются друг к другу с помощью специализированных клеточных соединений, называемых интеркалированными дисками. В интеркалированных дисках есть как якорные, так и щелевые соединения. Присоединенные клетки образуют длинные разветвленные волокна сердечной мышцы, которые действуют как синцитий, позволяя клеткам синхронизировать свои действия.Сердечная мышца перекачивает кровь по телу и находится под непроизвольным контролем.
Гладкая мышца Сокращение тканей отвечает за непроизвольные движения внутренних органов. Он образует сократительный компонент пищеварительной, мочевыделительной и репродуктивной систем, а также дыхательных путей и кровеносных сосудов. Каждая клетка имеет форму веретена с одним ядром и без видимых полосок (Рисунок 4.4.1 — Мышечная ткань).
Рисунок 4.4.1 — Мышечная ткань: (a) Клетки скелетных мышц имеют выраженную исчерченность и ядра по периферии.(б) Гладкомышечные клетки имеют одно ядро и не имеют видимых полос. (c) Клетки сердечной мышцы имеют поперечно-полосатую форму и одно ядро. Сверху: LM × 1600, LM × 1600, LM × 1600. (Микрофотографии предоставлены Медицинской школой Риджентс Мичиганского университета © 2012)
Внешний веб-сайт
Посмотрите это видео, чтобы узнать больше о мышечной ткани. Глядя в микроскоп, как можно отличить ткань скелетных мышц от гладких мышц?
Обзор главы
Три типа мышечных клеток: скелетные, сердечные и гладкие.Их морфология соответствует их специфическим функциям в организме. Скелетная мышца является произвольной и реагирует на сознательные раздражители. Клетки полосатые и многоядерные, выглядят как длинные неразветвленные цилиндры. Сердечная мышца непроизвольна и находится только в сердце. Каждая клетка имеет одно ядро и соединяется друг с другом, образуя длинные волокна. Клетки прикреплены друг к другу на вставных дисках. Клетки связаны между собой физически и электрохимически, чтобы действовать как синцитий.Клетки сердечной мышцы сокращаются автономно и непроизвольно. Гладкая мышца непроизвольна. Каждая клетка представляет собой веретенообразное волокно и содержит одно ядро. Никаких полос не видно, потому что актиновые и миозиновые филаменты не совпадают в цитоплазме.
Вопросы по интерактивной ссылке
Посмотрите это видео, чтобы узнать больше о мышечной ткани. Глядя в микроскоп, как можно отличить ткань скелетных мышц от гладких мышц?
Клетки скелетных мышц поперечно-полосатые.
Вопросы о критическом мышлении
Вы наблюдаете, как клетки в чашке спонтанно сокращаются.